МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
Розділ 3. РІСТ І РОЗМНОЖЕННЯ БАКТЕРІЙ
Поділ бактеріальної клітини. Клітинний цикл
Скоординований синтез мікроорганізмом компонентів клітини зумовлює її ріст, розмноження і в кінцевому результаті - утворення культури або колонії мікроорганізмів.
Для мікроорганізмів, як і для інших живих істот, характерні ріст і розмноження. Під ростом клітини мають на увазі узгоджене збільшення кількості всіх хімічних компонентів (наприклад, білка, ДНК, РНК), що супроводжується збільшенням розмірів і маси клітини. Ріст клітини не є безмежним. Досягнувши певних розмірів, клітина припиняє ріст і починає розмножуватися.
Розмноження - це збільшення кількості клітин мікроорганізмів у популяції. Проміжок від виникнення клітини до її поділу називають часом генерації або онтогенезом. У природі щодо цього спостерігається (за незначним винятком) певна закономірність: чим менший організм, тим швидше в нього з'являється нове покоління. Так, клітини E. coli діляться через кожні 20 хв. Коли б ніщо не перешкоджало їхньому поділу, то за добу виникло б стільки клітин, що з них можна було б побудувати піраміду з основою 1 км2 і заввишки 1 000 м.
Установлено певний взаємозв'язок між швидкістю росту клітини, її середньою масою і станом хромосоми. Перша подія, яка приводить до поділу клітини, - це ініціація реплікації хромосоми. У нормальних умовах цей процес взаємопов'язаний з масою клітини. Швидкість поділу бактерій залежить від наявності поживних речовин у середовищі, в якому ростуть клітини. Якщо не вистачає будь-яких важливих субстратів, знижується швидкість ініціації реплікації, але швидкість самої реплікації не змінюється.
Тривалість реплікації в більшості прокаріотичних клітин постійна (близько 40 хв). Якщо подвоєння ДНК розпочалося, воно триває до завершення незалежно від будь-яких змін у складі поживного середовища, які сталися за цей час. Час, який передує ініціації реплікації ДНК, непостійний (від 20 до 220 хв). Тривалий цикл має місце тоді, коли великий проміжок часу до початку ініціації. У цьому випадку після початку реплікації нові точки ініціації в хромосомі не виникають до того часу, поки не відбудуться всі події клітинного циклу. Час, необхідний для появи нових точок ініціації, непостійний, і переважно від нього залежить повна тривалість клітинного циклу. Короткий цикл спостерігається тоді, коли нові точки ініціації виникають до повного завершення циклу реплікації. Таким чином, ще до завершення поділу клітини розпочинається наступний цикл реплікації.
Ініціація реплікації здійснюється за допомогою специфічних білків. Якщо клітини помістити в середовище без відповідного ключового субстрату, синтез білка відразу ж припиняється. Хоча реплікація ДНК і поділ клітини тривають, новий цикл реплікації не розпочинається до того часу, поки знову не розпочнеться синтез білка. Звідси виходить, що ініціатори - це білки, які взаємодіють з ДНК і переводять її в такий стан, у якому вона здатна зв'язувати ДНК-полімеразу.
Реплікація ДНК розпочинається з точки прикріплення кільцевої хромосоми до ЦПМ, де локалізується ферментативний апарат, що відповідає за реплікацію. Реплікація, яка почалася в точці прикріплення, йде у двох протилежних напрямках, утворюючи характерну для кільцевої хромосоми проміжну структуру. У зоні реплікації (реплікацій- ній вилці) на невеликій ділянці відбувається розрив водневих зв'язків, які забезпечують підтримку двонитчастої структури ДНК.
На підготовлених таким чином однонитчастих ділянках, які служать матрицями, розпочинається
синтез комплементарних ниток ДНК (рис. 3.1).
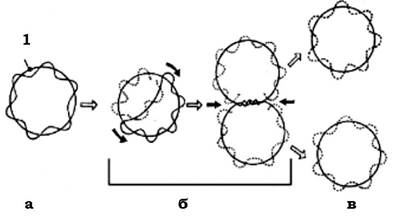
Рис. 3.1. Схематичне зображення реплікації кільцевої бактеріальної хромосоми:
а - материнська молекула ДНК; б - проміжні реплікативні форми; в - дочірні молекули ДНК після процесу реплікації і розходження:
1 - точка початку реплікації (чорними стрілками показано напрямок реплікації)
Після реплікації молекули ДНК залишаються зв'язаними з ЦПМ, яка починає рости між точками зв'язування ДНК і тим самим віддаляє їх, ніби розносячи в різні ділянки клітини. Це приводить до розділення (сегрегації) дочірніх молекул ДНК і оформлення відокремлених хромосом (рис. 3.2). Далі формується поперечна перетинка клітини.
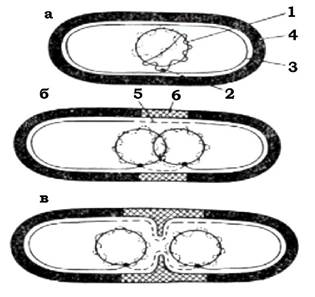
Рис. 3.2. Схематичне зображення механізму розподілу бактеріальних хромосом:
а - бактеріальна клітина містить частково репліковану хромосому, яка прикріплена до мембрани в точці реплікації; б - реплікація хромосоми завершена (у бактеріальній клітині дві хромосоми, кожна з яких прикріплена до ЦПМ; показано синтез клітинної стінки
1 ЦПМ); в - подальший синтез цитоплазматичної мембрани і клітинної стінки приводить до розділення дочірніх хромосом. Показано початок поділу клітини шляхом утворення поперечної перегородки: 1 - ДНК; 2 - прикріплення хромосоми до цитоплазматичної мембрани; 3 - ЦПМ; 4 - клітинна стінка; 5 - синтезована ділянка цитоплазматичної мембрани; 6 - новий матеріал клітинної стінки
За Р. Віттенбарі і С. Дау, формування поперечної перетинки та клітинної стінки в різних груп мікроорганізмів відбувається по-різному. Воно залежить від існування певних типів клітинного циклу:
- мономорфного, при якому в нормальних умовах утворюється лише один певний морфологічний тип клітин;
- диморфного, коли під час поділу утворюються дві клітини, які відрізняються між собою за формою, розмірами або мають інші ознаки відмінності;
- поліморфного, характерного для бактерій, здатних залежно від особливостей умов перебування утворювати два або більше морфологічних типів клітин.
Кожен із таких типів характеризується певними (нормальними) особливостями клітинного циклу.
Існування постійних типів поділу в мікроорганізмів свідчить про те, що він забезпечується спадковим матеріалом клітини.
Мономорфний клітинний цикл виявляється в більшості бактерій. Типовим представником бактерій з мономорфним клітинним циклом є кишкова паличка. У процесі росту клітини E. coli має місце лінійне неперервне збільшення її об'єму і поверхні. Перед поділом клітини стрибкоподібно збільшується швидкість її росту, і такий ріст можна характеризувати як білінійний.
Питання про механізм розходження дочірніх клітин грацилікутних бактерій залишається дискусійним. Так, деякі автори описують синхронний ріст усіх елементів оболонки. При цьому клітини діляться перетяжкою без утворення клітинної перетинки. Інші автори вважають, що перетинка формується з цитоплазматичної мембрани і шару муреїну.
У грампозитивних бактерій ріст міжклітинної оболонки можна простежити більш-менш чітко. Щоправда, механізми регуляції клітинного поділу вивчені недостатньо.
Характер росту і поділ фірмікутних коків, які належать до різних груп, дещо відрізняються. Ріст завжди йде вздовж осі симетрії клітини, а поділ - у площині, перпендикулярній до цієї осі. У стрептококів її напрямок стабільний, а це означає, що вони діляться в одній площині. У стафілококів поділ відбувається у двох взаємно перпендикулярних площинах, вісь симетрії після кожного поділу перемішується на 900. Поділ клітини у трьох взаємно перпендикулярних площинах, при відповідних переміщеннях осі симетрії, приводить до утворення пакетів.
З ростом клітини і збільшенням площі периферійної стінки зменшується периметр клітинної перетинки. Пропорційно до його зменшення відбувається вростання перетинки всередину клітини, що супроводжується зменшенням отвору, який з'єднує дочірні клітини. Площа поверхні перетинки майже не змінюється до її повного замикання. Поряд зі збільшенням площі клітинної стінки потовщується перетинка і нова частина периферійної стінки.
Клітинний поділ стафілококів відрізняється деякими специфічними особливостями (рис. 3.3).
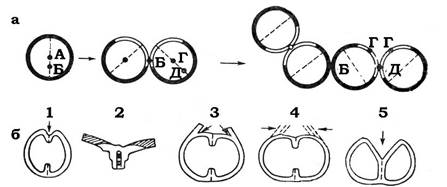
Рис. 3.3. Схема поділу клітини Staphylococcus aureus:
а - геометрія поділу: пунктирними лініями позначені переріз площини поділу; темним - частина оболонки, отримана від материнської клітини; світлим - заново синтезована частина оболонки; А-Д - точки на клітинній перетинці; б - механізм клітинного поділу: 1 - ініціація поділу; 2 - поперечний розріз зони ініціації (заштриховані ділянки материнської оболонки); 3 - дія злущувальної літичної системи; 4 - дія дезінтегруючої літичної системи; 5 - розщеплювальної літичної системи. Пунктиром позначена локалізація розщеплювальної літичної системи; стрілками - точки, в яких проявляється активність відповідних літичних систем
У стафілококів спочатку утворюється перетинка, розщеплення якої відбувається саме в момент поділу. Клітина, яка росте, залишається шароподібною або дещо овальною до самого моменту розходження дочірніх клітин. Перетинка розщеплюється швидко, але не повністю. Дочірні клітини набувають одразу округлої форми, але через те, що розщеплення не повне, клітини залишаються зв'язаними. Незважаючи на те, що площини послідовного поділу перпендикулярні одна до одної, нерозділені ділянки перегородки можуть бути розташовані випадковим чином, у результаті чого утворюються характерні гроноподібні скупчення клітин.
У бактерій роду Bacillus, на відміну від коків, процеси росту і поділу відбуваються неодночасно. Вони діляться за рахунок вростання і розщеплення перетинки. До того ж перетинка росте швидше, ніж розщеплюється. Новий муреїн синтезується і відкладається з внутрішнього боку оболонки. Старий муреїн відсовується до поверхні клітини, де, як передбачається, рівномірно розподіляється в міру її збільшення. При цьому виключно із заново синтезованого муреїну побудовані лише клітинні перетинки - полюси клітини після поділу та вузькі ділянки поверхні над перетинками.
Диморфний клітинний цикл - явище, яке спостерігається серед грамнегативних бактерій. Найкраще цей тип клітинного циклу досліджено в бактерій роду Caulobacter. Відбувається утворення двох типів клітин: рухомих (із джгутиками) і нерухомих (із стеблинками). І перші, і другі здатні ділитися, але їхні клітинні цикли відрізняються між собою (рис. 3.4). Нерухомі клітини зі стеблинками вважаються материнськими, а рухомі, які мають монотрихальний джгутик, - дочірніми.
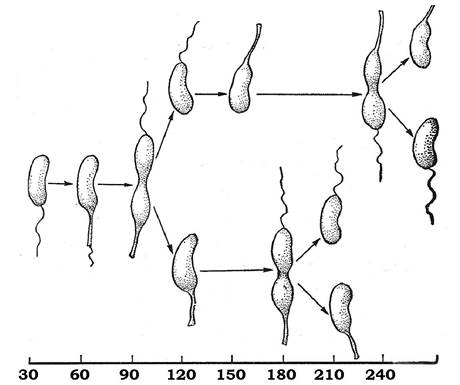
Рис. 3.4. Стадії життєвого циклу Caulobacter
Рухома клітина має декілька полярних пілей (фімбрій), які функціонують як рецептори для фагів, а також забезпечують адгезію на щільному субстраті. Рухомі клітини не здатні до поділу, поки в них не утвориться стеблинка, яка формується на протилежному джгутику полюсі. Утворення стеблинки супроводжується втратою джгутикового апарату і пілей.
Різновидом бінарного поділу є брунькування. Під час цього процесу на одному з полюсів материнської клітини утворюється невеличкий виріст (брунька), який збільшується під час росту. Коли брунька досягає розмірів материнської клітини, вона відділяється. Так, клітини Nitrobacter agilis і деяких фотосинтетичних бактерій роду Rhodopseudomonas діляться таким чином, що їхній ріст відбувається з одного полюса. У результаті утворюються дві нерівноцінні клітини. Дочірня клітина має джгутик, але він втрачається до початку ділення.
Розмноження типових представників мікоплазм, наприклад Mycoplasma mycoides, відбувається поділом надвоє: шляхом розпаду ниток і кілець на кокоподібні клітини і процесу, який нагадує брунькування (рис. 3.5). При цьому утворюються дрібні клітини, розміри яких коливаються в межах 0,125-0,150 мкм.
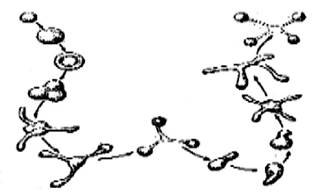
Рис. 3.5. Схема ділення мікоплазм у рідкому середовищі
Деякі представники роду Hyphomicrobium проявляють, залежно від умов культивування, здатність до поліморфного клітинного циклу. Так, під час вирощування їх на середовищі з метанолом утворюються гіфи й овальні бруньки (рис. 3.6), а з метиламіном - кістеподібні клітини, від яких відбруньковуються рухомі дочірні особини. Поліморфізм деяких штамів Hyphomicrobium спостерігається й на інших субстратах.
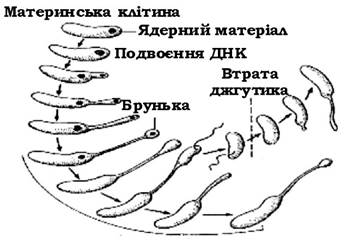
Рис. 3.6. Стадії життєвого циклу Hyphomicrobium
У деяких ціанобактерій описано розмноження шляхом множинного поділу, яке розпочинається зі збільшення розмірів вегетативної клітини яка з часом зазнає ряду швидких бінарних поділів, що відбуваються всередині додаткового фібрилярного шару материнської клітинної стінки. Це супроводжується утворенням дрібних клітин, які отримали назву баеоцитів (маленьких клітинок).
Кількість їх у різних видів коливається від 4 до 1 000. Вивільнення баеоцитів відбувається під час розриву материнської клітинної стінки (рис. 3.7), тобто в основі множинного поділу лежить принцип рівновеликого бінарного поділу, особливість якого полягає в тому, що ріст дочірніх клітин не відбувається: вони знову діляться.
Таким чином, у процесі поділу бактеріальної клітини визначаються як мономорфний, так і ди- та поліморфний цикли розвитку.
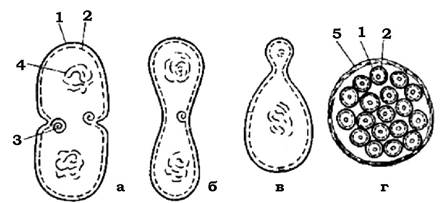
Рис. 3.7. Схематичне зображення способів поділу прокаріотичної клітини:
а - поділ шляхом утворення поперечної перегородки; б - поділ шляхом перетяжки; в - брунькування; г - множинний поділ: 1 - клітинна стінка; 2 - ЦПМ; 3 - мембранна структура; 4 - нуклеоїд; 5 - додатковий фібрилярний шар клітинної стінки
Спосіб поділу бактеріальної клітини не можна встановити при поверхневому розгляді препарату. Для цього необхідні тривалі спостереження за поведінкою живих клітин. Методи простого забарвлення також не дозволяють з необхідною точністю визначити, до якого типу належить цей поділ.
Як у лабораторних, так і у виробничих умовах одним із важливих завдань є отримання максимальної кількості продукту життєдіяльності мікроорганізму. У всіх випадках, особливо при виробництві мікробної біомаси, основним показником біотехнологічного процесу є інтенсивність росту і розмноження клітин. Ріст і розмноження мікроорганізмів - не постійні величини, вони залежать від поживної цінності середовища, фізико-хімічних умов культивування, фізіологічного стану (віку) культури. Поняття про вік мікроорганізмів цілком визначене в розумінні онтогенетичного циклу, але досить відносне в часі. Перебування в молодому віці і процес старіння мікробної популяції залежать від умов культивування. Основна причина старіння - накопичення продуктів метаболізму і виснаження поживного середовища. При постійному відведенні продуктів метаболізму і надходженні поживних речовин мікробні клітини можуть тривалий час перебувати молодими. Використовуючи такі технологічні прийоми, можна створити умови, за яких мікробна популяція буде перебувати в стані постійної активної життєдіяльності.
В ідеальних умовах, при постійному відведенні продуктів метаболізму і надходженні поживних речовин, можна спостерігати нескінченний ріст і розмноження клітин мікробної популяції. Знаючи час генерації даного мікроорганізму (g) і початкову кількість клітин (Х0), можна визначити їхню кількість у будь-який момент часу культивування (t):
Х =Х0 ·2n,
де Х - кількість клітин у момент t; Х0 - початкова кількість клітин у момент t0; n - кількість генерацій:
n = t / g ,
де t - час культивування; g - час генерації (подвоєння кількості) клітин.
Тобто ріст мікробної популяції визначається показником ступеня або кількістю поділів (генерацій), яка стоїть в експоненті (показнику ступеня). Тому дана залежність є експоненціальною.
Якщо уявити, що час генерації для Escherichia coli становить 20 хв, то з однієї єдиної клітини протягом 24 год можна отримати кількість клітин, яка становитиме Х = 1 · 272 = 4,72 · 1021
Інша картина спостерігається в дійсності, особливо при періодичному культивуванні. Періодичною називають систему, в якій після інокуляції (посіву мікроорганізму) середовища відповідного складу не відбувається ні внесення стерильних порцій поживного середовища, ні вилучення будь-яких продуктів метаболізму.