МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
Розділ 3. РІСТ І РОЗМНОЖЕННЯ БАКТЕРІЙ
ЗАКОНОМІРНІСТЬ РОЗВИТКУ МІКРОБНОЇ ПОПУЛЯЦІЇ В ПЕРІОДИЧНІЙ СИСТЕМІ
Питання росту та розмноження мікроорганізмів вивчав Ж. Моно (1942). За Моно, розмноження популяції бактерій відбувається таким чином: при засіві бактерій на свіже поживне середовище вони спочатку пристосовуються до нових умов і певний час не розмножуються (lag-фаза); потім починається фаза швидкого розмноження з логарифмічною залежністю кількості клітин від часу вирощування (log- фаза); поступово поживні речовини в середовищі вичерпуються, і бактеріальна популяція вступає в стаціонарну фазу; останньою відбувається фаза відмирання та лізису клітин (рис. 3.8).
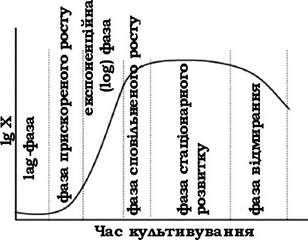
Рис. 3.8. Ідеалізована крива росту бактеріальної популяції в періодичній системі культивування
Популяція (франц. population - населення) - це сукупність бактерій одного виду (чиста культура) або різних видів (змішані культури), які розвиваються в обмеженому просторі. У бактеріальній популяції постійно відбувається ріст бактерій, їх розмноження та відмирання. При спостереженні за розвитком бактеріальної популяції увагу звертають, перш за все, на кількість клітин (або біомасу) в одиниці об'єму та їхню життєздатність. З цією метою використовують прямі та непрямі методи. За допомогою прямих методів підраховують загальну кількість клітин у популяції або визначають їхню сумарну масу в певному об'ємі поживного середовища. Кількість клітин підраховують у лічильних камерах під мікроскопом із застосуванням електронного лічильника, шляхом підрахунку клітин на мембранних фільтрах та ін. Для визначення лише кількості життєздатних клітин використовують посів бактеріальної суспензії на щільні поживні середовища з наступним підрахунком колоній, які виросли. При цьому вважається, що кожна колонія виросла з однієї клітини.
Потрапляючи в нове середовище, клітини мікроорганізмів повинні адаптуватися до його умов (lag-фаза, або фаза затримки росту). У результаті різкої зміни умов ферментні системи мікроорганізмів деякий час перебувають у пригніченому стані. Тривалість цієї фази залежить як від зовнішніх умов, так і від віку бактерій та їхньої видової специфічності. У тих випадках, коли джерело енергії і вуглецю в новому середовищі відрізняються від тих, які були при попередньому культивуванні, адаптація до нових умов може бути пов'язана із синтезом нових ферментів, які раніше не були потрібні. Синтез нових ферментів може індукуватися новими субстратами середовища. Зміни у складі бактеріальної клітини під час lag-фази позначаються насамперед на вмісті РНК, який збільшується у 8-12 разів. Таке збільшення кількості РНК свідчить про її участь у синтезі ферментів.
З часом мікробні клітини починають синтезувати ферменти, необхідні для асиміляції компонентів існуючого середовища, збільшується середній об'єм клітин і бактеріальна популяція набуває здатності до розмноження. Розпочинається фаза прискореного росту (див. рис. 3.8).
Молоді клітини мають більшу спорідненість до основних барвників - їхня забарвленість на препаратах інтенсивніша, ніж у старих клітин. На підставі цього було зроблено припущення, що ізоелектрична точка їхньої протоплазми зсунута в бік низьких значень pH. Поглинання кисню, утворення тепла, СО2 й аміаку вказує на те, що обмін речовин у молодій клітині відбувається більш інтенсивно. Молоді клітини краще, ніж старі пристосовуються до нового поживного середовища, але вони менш стійкі до дії зовнішніх факторів (фенолу, нагрівання, охолодження, солей тощо).
Потім клітини інтенсивно розмножуються, культура мікроорганізмів переходить в експоненційну фазу, або як її називають, фазу логарифмічного розвитку. Вона характеризується постійною максимальною швидкістю поділу клітин, яка залежить від виду мікроорганізму й умов середовища. Кількість мікробних клітин у цей період збільшується в геометричній прогресії за експоненційним законом (рис. 3.9).
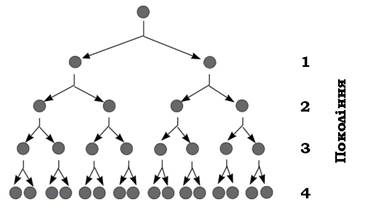
Рис. 3.9. Схема розмноження бактеріальної клітини
Розмір клітин і вміст білка у клітинах багатьох бактерій залишаються незмінними протягом усієї експоненційної фази, а процеси росту проходять збалансовано - подвоєння біомаси супроводжується подвоєнням вмісту білка, РНК, ДНК та ін. У таких умовах за ростом культури можна простежити, визначаючи один із цих показників.
Стаціонарна фаза починається тоді, коли кількість клітин перестає збільшуватися. Швидкість росту залежить від концентрації субстрату в середовищі.
Перехід від експоненційної фази до стаціонарної відбувається поступово. Швидкість росту може знижуватися не лише через зменшення поживного субстрату, але також через збільшення щільності бактеріальної популяції, зміну парціального тиску О2 або накопичення продуктів метаболізму. Усі ці фактори викликають перехід до стаціонарної фази. Настає рівновага, коли кількість клітин (біомаси) не збільшується і не зменшується, тому що кількість клітин, які відмирають, дорівнює кількості клітин, які утворилися в результаті розмноження.
З часом кількість відмерлих клітин починає домінувати над тими, що утворюються і крива росту мікробної популяції опускається вниз. Настає фаза відмирання, яка супроводжується лізисом мікробних клітин. Кількість життєздатних клітин може знижуватися експоненційно. Інколи клітини лізуються під дією власних ферментів (автоліз). Такий стан бактеріальної популяції зумовлюється зміною фізико-хімічних властивостей поживного середовища та іншими несприятливими чинниками.
Розвиток культур мікроорганізмів прийнято характеризувати деякими кількісними показниками, які використовуються на практиці. До таких показників відносять: величину (розмір) мікробної популяції; швидкість росту; час генерації та економічний коефіцієнт.
Величина розмір) мікробної популяції визначається її чисельністю або сумарною масою її особин, тобто розмір мікробної популяції можна подати як кількість клітин (або їх масу) у певному об’ємі культуральної рідини. При цьому мікробна популяція характеризується маленькою масою окремих особин (клітин) і великою їх кількістю в одиниці об’єму.
Валова швидкість росту - це збільшення кількості біомаси за одиницю часу:
![]()
Але це величина абсолютна, вона вказує лише на зміни загальної кількості біомаси в даних конкретних умовах і не може бути використана для порівняння виробництв різного масштабу та оцінки ефективності даного мікроорганізму. Більш реальне уявлення про ці характеристики дає питома швидкість росту, яка визначається як відношення валової швидкості росту до початкової біомаси:
![]()
Після інтегрування рівняння (2)
![]()
а після переводу в десятковий логарифм
![]()
де Х та Х0 - кількість клітин (або будь-якого компонента клітини) у момент часу t і t0 відповідно.
Знаючи Х0 та визначивши Х, можна визначити μ мікробної популяції, виходячи з рівняння (4). Наприклад, якщо в момент t популяція мала 104, а через 4 год (момент t0) - 108клітин, тоді
![]()
З рівняння (3) видно, що μ (t - t0) - це показник ступеня натурального логарифма, бо цей вираз одержують при логарифмуванні рівняння
![]()
де е - основа натурального логарифма.
Виходячи з рівняння (6), можна визначити кількість біомаси в будь-який період росту:
![]()
Кількість біомаси, яка накопичується в стаціонарній фазі, називають виходом або врожаєм (рис. 3.10). Він залежить від природи та кількості поживних речовин, які використовуються, а також умов культивування.
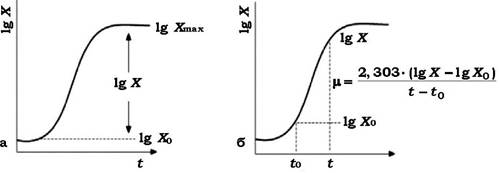
Рис. 3.10. Параметри росту:
а - врожай клітин, б - швидкість росту
Час генерації - це час подвоєння кількості клітин (або компонентів мікробної клітини). Цей параметр можна визначити з рівняння (3). При цьому виходять із того, що t - t0 = g, тоді Х = 2 Х0. У таких умовах рівняння (3) матиме вигляд
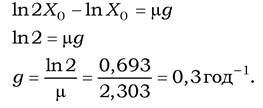
Швидкість розмноження (коефіцієнт розмноження) - це кількість поділів за одиницю часу:
![]()
Тривалість кожної генерації можна виразити як
![]()
Наведених кількісних показників, пов'язаних лише з ростом культур, недостатньо для повної характеристики мікробіологічного процесу. Необхідно враховувати також зміни, які відбуваються в середовищі в результаті життєдіяльності клітин, а саме зменшення концентрації поживних речовин та накопичення продуктів метаболізму.
Між питомою швидкістю росту і швидкістю асиміляції субстрату існує певна залежність
![]()
де μ mах - максимальна швидкість росту культури (константа росту), поріг, до якого прагне швидкість росту в міру підвищення концентрації субстрату, який асимілюється; S - концентрація субстрату, який асимілюється; K - константа насичення, що дорівнює концентрації поживного субстрату, при якій швидкість росту досягає половини граничної, тобто
![]()
Одним із факторів, який визначає швидкість росту культури, є накопичення продуктів метаболізму. Існує залежність між утворенням продуктів метаболізму і питомою швидкістю росту культури:
![]()
де μ0 - питома швидкість росту культури в даному середовищі за повної відсутності продуктів метаболізму; Р - фактична концентрація продуктів метаболізму; Kр - константа, що дорівнює концентрації продуктів обміну, при якій швидкість росту знижується вдвічі, тобто при![]()
Важливим показником розвитку культури мікроорганізмів є економічний коефіцієнт, який визначається як відношення приросту біомаси (врожаю, Х) до кількості витраченого субстрату (S), - вихід біомаси з одиниці субстрату:
![]()
Якщо врожай (у грамах) відносять до кількості молів асимільованого субстрату, то економічний коефіцієнт, який називається в даному випадку молярним економічним коефіцієнтом, позначають через Ym. Наприклад, YАТФ - це енергетичний коефіцієнт, який подається в грамах клітинної маси, отриманої при витраті 1 моля АТФ.
Наведена на рис. 3.8 крива росту мікробної популяції ідеалізована. У комплексному середовищі бактеріальні клітини часто використовують субстрати в певній послідовності. Наявність певних субстратів може супроводжуватися репресією синтезу ферментів, які беруть участь у метаболізмі інших поживних речовин. У цьому випадку ферменти, що каталізують метаболізм деяких речовин, починають діяти лише після того, як концентрація субстратів, які репресують їхній синтез, зменшиться в результаті асиміляції клітинами. Регуляція фізіології бактерій супроводжується зміною кривої росту і появою однієї або декількох перехідних стаціонарних фаз. Таку відповідь культури на зміни в складі середовища називають діауксією. Класичним прикладом діауксії може бути ріст Escherichia coli на середовищі, яке містить глюкозу і лактозу (рис. 3.11). Спочатку відбувається ріст культури за рахунок використання глюкози. Після засвоєння цього субстрату крива росту різко змінює свій нахил, а кількість клітин може зменшитися. Потім знову настає lag-фаза під час якої відбувається індукування нової ферментної системи, що буде каталізувати асиміляцію лактози, і накопичення біомаси продовжиться.
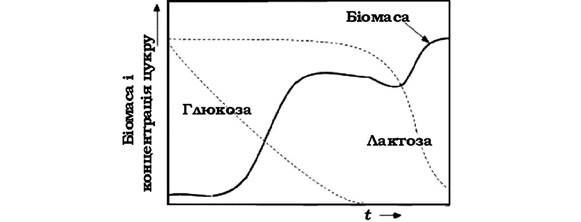
Рис. 3.11. Ідеалізована діауксія росту бактерій у періодичний системі культивування в середовищі, яке містить два субстрати (глюкоза і лактоза)
У процесі періодичного культивування виникають певні проблеми, пов'язані зі зміною морфології та фізіології культури, концентрації поживних речовин і продуктів метаболізму. Вплив цих факторів супроводжується зміною швидкості розвитку мікробної популяції, що завершується припиненням процесу культивування.