МИКРОБИОЛОГИЯ БИОЛОГИЯ ПРОКАРИОТОВ ТОМ I - А. В. ПИНЕВИЧ - 2006
ГЛАВА 5. ПРОКАРИОТЫ В СВЕТЕ КЛЕТОЧНОЙ ТЕОРИИ
С анатомической точки зрения основной жизненной единицей является клетка, в которой все проявления жизни и должны найти свое выражение в наиболее простой и ясной форме.
Ю. А. Филиппенко. Общедоступная биология (Петроград, 1923).
Живые системы так разнообразны в структурно-функциональном плане, что дать концептуальное определение жизни и одновременно с этим избежать редукционизма, когда общее подменяется частностями — невыполнимая задача.
Тем не менее, среди атрибутов жизни можно выбрать наиболее характерный, что делалось неоднократно с позиций соответственно термодинамики, информатики, органической химии и теории конкурентной эволюции.
Русский эволюционист и зоогеограф Л. С. Берг в 1922 г. определил жизнь на основе понятия целесообразности. По его мнению, живыми можно назвать физические тела, целесообразно отвечающие на внешнее раздражение.
Один из основоположников теоретической биологии, русский физиолог Э. С. Бауэр сформулировал в 1935 г. определение жизни с позиций термодинамики. Согласно ему, основой жизни служат неравновесные термодинамические процессы, и живые системы создают внутреннюю упорядоченность за счет увеличения неупорядоченности в абиотической среде, накапливая «отрицательную» энтропию.
Определение жизни с позиций информатики принадлежит одному из создателей квантовой теории, австрийскому физику Эрвину Шредингеру (Е. Schrödinger, Нобелевская премия по физике, 1933 г.). В лекциях, прочитанных в 1943 г., он говорил, что главным свойством жизни является линейная анизотропия органических макромолекул, позволяющая им играть роль носителей наследственной информации.
Известный английский химик и философ Джон Бернал (J. Bernal) в своих теоретических работах 1960-х годов рассматривал жизнь с позиций органической химии — как самосохраняющуюся сеть сопряженных реакций, которые осуществляются между органическими молекулами при участии эндогенных органических катализаторов.
В экзобиологической программе NASA 1984 г. жизнь определяется в свете теории конкурентной эволюции — как автономная физико-химическая система, подчиняющаяся принципу селекционизма, т. е. естественному отбору по Дарвину.
Во всех приведенных определениях не учитывается то, что жизнь представляет собой конкретный этап химической эволюции Вселенной. Уникальными особенностями жизни являются:
— существование на основе клеток и совокупностей особей;
— адаптивное взаимодействие с окружающей средой;
— индивидуальное развитие, или онтогенез;
— историческое развитие, или филогения.
Решающий вклад в понимании информационных основ жизни сделали американский вирусолог Джеймс Уотсон (J. Watson), английский молекулярный биолог Френсис Крик (F. Crick) и английский биофизик Морис Уилкинс (М. Wilkins), которые установили, что гетерополинуклеотиды обладают способностью к матричному копированию (Нобелевская премия по физиологии и медицине, 1962 г.). Дальнейшие исследования привели к расшифровке универсального генетического кода. Они показали, что хранящие информацию полинуклеотиды не только автокопируются (реплицируются), но и в промежутке между этим служат источником команд для образования белков и надмолекулярных систем, которые многоэтапным путем обеспечивают все тот же процесс матричного копирования.
Последовательность событий складывается из транскрипции (синтеза РНК на матрице ДНК), трансляции (синтеза белка на матрице РНК), посттрансляционного процессинга и, наконец, функциональной самореализации белка в субклеточных системах.
Структура полинуклеотидных матриц такова, что они не только реплицируются, но и способны:
— к репарации, т. е. восстановлению повреждений;
— к мутации, т. е. наследуемому локальному изменению структуры в результате неадекватной репарации;
— к рекомбинации, т. е. передислокации эндогенных блоков или включению экзогенных блоков;
— к амплификации, т. е. кратному увеличению числа имеющихся генетических элементов.
Все эти преобразования протекают на конкурентной основе, с общей тенденцией к повышению
гетерогенности матриц и увеличению объема содержащейся в них информации.
Химической основой этого процесса служит реакция гетерополимеризации. Ее катализируют белки — продукты матричного копирования на альтернативный полипетидный носитель.
Для копирования матриц требуются биосинтетические системы. Они состоят из ферментносубстратных комплексов и нуждаются в энергии. Ферменты, субстраты и энергия служат также для защиты геномов от повреждений, искажающих или блокирующих процесс матричного копирования. В частности, ДНК должна быть структурно стабилизирована и, при необходимости, подвергаться репарации.
Для полимеразных реакций обоих типов необходимо сконцентрировать органическое вещество и биологически доступную энергию.
На начальном этапе эволюции полимеразные реакции проходили в среде, обогащенной продуктами абиотического синтеза (первичном «бульоне»). По мере истощения этих запасов среда становилась все более олиготрофной. Поэтому примитивные живые системы начали концентрировать конструктивные субстраты и энергоносители, а затем активно их преобразовывать. В современных природных условиях матричное копирование может происходить только в изолированном пространстве — компартменте, внутренняя среда которого находится в состоянии устойчивого неравновесия с окружающей средой. Для того, чтобы эта ситуация воспроизводилась, компартмент должен быть квази-стационарным, т. е. быть открытой системой. Клетка, окруженная полупроницаемой мембраной, отвечает этому требованию. Она представляет собой химический реактор, в котором протекает нуклеотид-полимеразная реакция. Копироваться может как эндогенная матрица (клеточный геном), так и экзогенная матрица (плазмида или геном паразитического организма, не имеющего клеточного строения).
Копирование матриц — потенциально бесконечный процесс, и в этом смысле органическая жизнь исключительно «заразна», т. е. способна к неограниченной пространственно-временной экспансии за счет освоения всех доступных ресурсов энергии и субстрата.
Учитывая сказанное, мы можем дать еще одно определение органической жизни: органическая жизнь — это потенциально неограниченная экспансия полинуклеотидных матриц, копирование которых происходит в клетках, или мультивариантных мембранных компартментах, способных к самовоспроизведению.
Оно относится не только к живым существам, обладающим клеточным строением, но и к неклеточным живым существам — вирусам, которые реплицируются внутри клеток-хозяев. Происхождение вирусов неизвестно, и они могут соответствовать доклеточной, вторично упрощенной клеточной или независимо возникшей неклеточной форме жизни. В геноме вируса закодированы только те системы, которые обеспечивают его репликацию после внедрения в клетку, но не кодируют функциональные структуры самой клетки. В отличие от вирусов, клетки представляют собой автономную форму жизни, хотя в известном смысле вирусы более совершенны.
В приведенном определении жизни наряду с метаболическим и генетическим аспектами ведущая роль отводится пространственной организации копирования полинуклеотидных матриц. Именно это позволяет считать цитоархитектонику центральным аспектом биологии клетки.
В отношении цитоархитектоники живые организмы образуют не сплошной ряд, а дискретный набор морфотипов. Классифицируя последние, мы создаем фенотипическую мегасистему, а насколько она объективна, скажут современные представления о глобальной филогении.
5.1. Современное определение клетки. Новое содержание терминов «Прокариоты» и «Эукариоты»
Учение о двух морфотипах, эукариотном и прокариотном, исходит в первую очередь из того, имеется ли в клетке морфологически оформленное ядро. Однако ядро является не единственным признаком, по которому прокариоты и эукариоты отличаются друг от друга.
Еще в 1970-е годы Роджер Стэниер, Лин Маргелис и Додсон (Е.Dodson) независимо друг от друга пришли к выводу, что при сравнении прокариотов с эукариотами необходимо учитывать не один, а множество критериев — от молекулярных до морфологических. Они пытались ранжировать признаки клетки на главные и второстепенные, универсальные и неуниверсальные. Однако такой подход имеет очень существенный недостаток — вместо интегральной характеристики морфотипа составляется перечень признаков, соответствующих разным уровням клеточной организации. Альтернативный подход заключается в том, чтобы из совокупности признаков морфотипа выбрать важнейший.
В начале 1980-х годов Томас Кавалье-Смит пришел к следующему принципиальному заключению: эукариоты принципиально отличаются от прокариотов общим планом строения мембранной системы, т. е. способом компартментализации клетки.
Естественно, что у ядерных клеток не принимаются во внимание митохондрии и пластиды. Эти органеллы, или «ксеносомы» (греч. ksenos — чужой и soma — тело) произошли от бактерий. По типу строения они остаются клетками, несмотря на то, что их генетическая, структурная и физиологическая автономия во многом утрачена.
Главным признаком эукариотной клетки, ее «маркером», служит ядерная оболочка. Некоторые свойства этой мембранной структуры уникальны, а ее функциональную роль трудно недооценить. Однако если взглянуть на нее под более широким углом зрения, то выясняется, что речь идет об обособленном и функционально дифференцированном участке эндомембранной системы. Проще говоря, специфика эукариотов определяется не ядерной оболочкой, а общими свойствами их эндомембранной системы.
Прежде всего, эукариоты не не могут существовать без эндомембран, которые сохраняются в клетках этого морфотипа независимо от уровня их эволюционного развития и особенностей онтогенеза.
Кроме того, для эукариотов характерен специальный тип функционального взаимодействия между эндомембранной системой и цитоплазматической мембраной — они структурно разобщены и взаимодействуют друг с другом с помощью везикулярного транспортного аппарата.
Часть рибосом эукариотов прикрепляется к мембранам эндоплазматического ретикулума, которые растут посредством встраивания белков и липидов, в то время как рост цитоплазматической мембраны происходит в результате слияния с ней везикул, отпочковывающихся от аппарата Гольджи. Напротив, в прокариотной клетке рибосомы могут прикрепляться прямо к цитоплазматической мембране.
Таким образом, принципиальное различие между эукариотами и прокариотами заключается в том, что эукариоты всегда содержат эндомембраны. Зато прокариотам иметь эндомембраны не обязательно. Если они у них есть, то взаимодействуют с цитоплазматической мембраной постоянно или только временно, причем это всегда прямое взаимодействие, без участия везикулярного транспортного аппарата.
Разные участки эндомембранной системы эукариотов различаются по морфологии и выполняют конкретные функции, связанные с транспортом и конструктивным метаболизмом. В отличие от этого, эндомембраны прокариотов — однообразного строения и осуществляют энергетический метаболизм.
Итак, эукариоты и прокариоты различаются типом организации мембранной системы, хотя следует еще раз напомнить о том, что эукариоты монофилетичны, а прокариоты дифилетичны, и под названием «прокариоты» выступают представители двух глобальных доменов — Bacteria и Archaea. Их разделяет примерно такая же эволюционная дистанция, которая каждого по отдельности разделяет с эукариотами.
Выходит, что дихотомическая классификация устарела. Означает ли это, что также устарели термины «прокариотная клетка» и «эукариотная клетка»? Чтобы ответить на этот вопрос, нам нужно уточнить, что мы подразумеваем под клеткой.
Клетка представляет собой микрокомпартмент для хранения и воспроизведения генетической информации. Его важнейшими и незаменимыми компонентами являются:
— ограничивающая мембрана;
— геном, в котором закодирована индивидуальная белоксинтезирующая система;
— клонированная популяция рибосом.
Всеми перечисленными атрибутами обладает элементарная клетка, или «монада».
5.1.1. Монады и химеры
Поскольку понятия «монады» и «химеры» имеют концептуальное значение для цитологии, остановимся на них более подробно.
Начиная с античности, философский термин «монада» (греч. monos — единица, единое) служит для обозначения основополагающего и неделимого элемента бытия. Слово «монада» и производные от него понятия в разное время использовались как биологические термины. Чаще всего монадами называли такой тип биологической организации, когда клетка представляет собой отдельный организм, что приблизительно соответствует содержанию, которое теперь вкладывается в термин «протисты». В некоторых случаях слово «монада» становилось частью таксономического названия, например, бактерии Azomonas, Hyphomonas и Pseudomonas, археот Natronomonas, зеленая водоросль Chlamydomonas и т. д. Гипотетическим монадам приписывали ключевую роль в эволюции («монеры» Эрнста Геккеля).
Фрэнсис Тэйлор использовал в 1982 г. термин «монада» в общецитологическом смысле для того, чтобы провести различие между «монадами» (собственно клетками) и «химерами» (эндосимбиотическими системами).
Клетка-монада является элементарной единицей жизни. Атрибуты монады, или набор структур, которые необходимы в онтогенезе и для размножения, следующие:
— цитоплазматическая мембрана, или мембранный барьер, окружающий цитоплазматический компартмент;
— геном, или индивидуальная кодирующая система на основе компактизирован- ной двухцепочечной ДНК;
— клонированная популяция рибосом, или универсальных органелл трансляции.
Четвертым атрибутом монады служит цитоплазма, но она аморфна и поэтому не рассматривается в качестве элемента клеточной архитектуры. Наконец, можно было бы назвать пятый атрибут монады, цитоскелет. Однако у разных монад он выполняет разные функции, не обязательно связанные с клеточной архитектурой и ключевыми процессами жизнедеятельности.
Таким образом, исходя из приципа: «Одна цитоплазматическая мембрана-один геном-одна клонированная популяция рибосом», мы можем говорить о двух типах монад. Это прокариотная монада и эукариотная монада.
Всеми атрибутами прокариотной монады обладают и эндосимбиотические органеллы бактериального происхождения — митохондрии и «простые» пластиды. Они имеют мембрану, окружающую матрикс (в случае митохондрий) или строму (в случае «простых» пластид), хромосому и набор 70S рибосом.
Монады, обладающие собственным мембранным барьером, индивидуальным геномом и клонированной популяцией рибосом, могут быть только двух типов — прокариотная монада и эукариотная монада.
Монады способны образовывать группы клеток — полимерные ассоциации монад. Для их обозначения Тейлор предложил термин «полимонада» (англ. polymonad).
Полимонады могут состоять из монад с гомологичными геномами. Примером «гомологичной» полимонады, состоящей из фототрофных особей, служат бактерии рода Pleurocapsa.Примером «гомологичной» полимонады, состоящей из хемотроф- ных особей, служат бактерии рода Streptococcus (рис. 3).
Рис. 3. Клетки-монады, эндосимбиотические системы-химеры и полимерные ассоциации монад или химер. N — ядро; n — нуклеоморф (рудимент ядра).
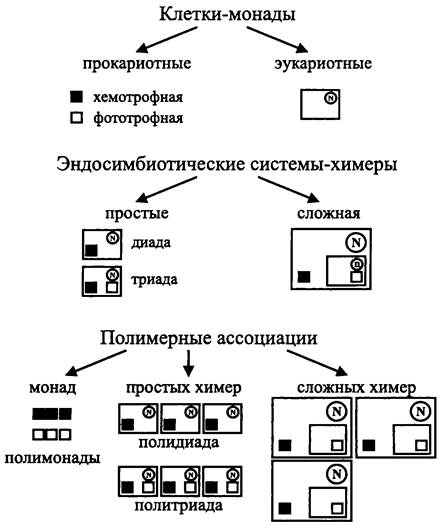
Кроме того, полимонады могут состоять из монад с гетерологичными геномами. Такие ассоциации называются консорциумами (лат. consortium — общность). Примерами может служить консорциум, образованный сульфатредуцирующими гетеротрофными бактериями и зелеными фототрофными бактериями, а также консорциум, образованный гипертермофильными археями Igniococcus sp. и «Nanoarchaeum equitans».
Эндосимбиотические системы представляют собой принципиально иной тип межклеточной ассоциации. В этом случае прокариотная монада поселяется внутри эукариотной монады. В результате они становятся настолько взаимозависимыми, что уже не могут существовать друг без друга. Именно таким путем в ходе эволюции возникли митохондрии (потомки хемотрофных прокариотных монад) и «простые» пластиды (потомки фототрофных прокариотных монад).
Для эндосимбиотической ассоциации прокариотных монад с эукариотной монадой Тейлор предложил термин «простая химера» (англ. simple chimera; греч. chimaira — Химера, мифологический монстрозный гибрид льва, козы и змеи). В зависимости от состава партнеров различаются два типа простых химер.
Простая химера, образованная эукариотной монадой и хемотофной прокариотной монадой, называется «диадой» (англ. diad; греч. dio —два). В свою очередь, простая химера, образованная эукариотной монадой, хемотрофной монадой и фототрофной монадой, называется «триадой» (англ. triad; греч. tris — три). Ассоциации диад и триад называются «полидиадами» (англ. polydiad) и «политриадами» (англ. polytriad).
Большинство представителей царства Animalia, имеющие митохондрии, — это полидиады. В свою очередь, большинство представителей царства Plantae, имеющие митохондрии наряду с «простыми» пластидами — это политриады.
Количественную основу современного биоразнообразия составляют животные (1032 тыс. видов, из них только 751 тыс. видов насекомых), высшие растения (248 тыс. видов), грибы (69 тыс. видов), нефотосинтезирующие протисты (30 тыс.
видов) и водоросли (27 тыс. видов). Эти организмы являются потомками эндо- симбиотических ассоциаций между фаготрофными эукариотными клетками (домен Еuсаrуа) и представителями двух разных фил домена Bacteria — BXII Proteobacteria и ВХ Cyanobacteria. В данном случае монады играют роль соответственно клеток- хозяев, митохондрий и «простых» пластид.
Более сложный вариант эндосимбиотической ассоциации возникает, когда одна химера поселяется внутри другой. Такая эндосимбиотическая система назвается «сложной химерой» (англ. complex chimera). Примеры сложных химер — многие фототрофные протисты, в частности представители классов Cryptophyceae и Chlorarachniophyceae, сохранившие рудимент ядра (нуклеоморф). В настоящее время установлено, что «сложные» пластиды этих микроорганизмов произошли не от бактерий, а от водорослей-триад, которые имели митохондрии и простые пластиды. Путем эндоцитоза такие триады были захвачены фаготрофными диадами и в результате эволюции межклеточной ассоциации превратились в фототрофные органеллы.
Сложные химеры также могут образовывать полимерные ассоциации. Примером служат бурые, золотистые и желто-зеленые водоросли, которые потеряли нуклеоморф, но сохранили рудиментарные «эукариотные» мембраны вокруг «простой» пластиды.
Возвращаясь к монадам, необходимо подчеркнуть, что учтенное биоразнообразие этих организмов (5,2 тыс. видов бактерий и архей, а также несколько десятков видов «амитохондриальных» протистов) ничтожно мало по сравнению с биоразнообразием химер (1406 тыс. видов).
5.1.2. Прокариоты и эукариоты как два типа монад
На основе современных представлений о свойствах клетки и ее филогении прокариотам можно дать следующее определение: прокариоты — это неядерные монады, из которых состоят филогенетические домены Bacteria и Archaea.
В свою очередь, эукариотам можно дать следующее определение: эукариоты — это ядерные монады, из которых состоит филогенетический домен Еuсаrуа.
Представители филогенетического домена Еuсаrуа обладают эукариотным морфотипом. Его характерной чертой является уникальный мембранный аппарат, который подразделяется на две обособленные части — цитоплазматическую мембрану и эндомембраны.
Эндомембраны эукариотов представляют собой совокупность компартментов, сообщающихся между собой при помощи системы везикулярного транспорта. Эндомембраны взаимодействуют с цитоплазматической мембраной при помощи везикулярной челночной системы, которая осуществляет цитоз, или «транспорт в упаковке». Он происходит из цитоплазмы во внеклеточную среду и в противоположном направлении — из внеклеточной среды в цитоплазму. Кроме того, с помощью везикулярной челночной системы переносится материал, из которого состоит цитоплазматическая мембрана.
Эндомембраны эукариотов имеют второе название — «вакуом» (лат. vacuola — пузырек и omnis — совокупный). В 1980-е годы его ввел бельгийский биохимик Кристиан де Дюв (С. de Duve, Нобелевская премия по физиологии и медицине, 1974 г.). Этим термином обозначается система топологически замкнутых мембранных микрокомпартментов, или вакуолей в широком смысле слова.
В состав вакуома входят ядерная оболочка, аппарат Гольджи, транспортные везикулы, эндоплазматический ретикулум, микросомы и т. д. Функциями вакуома являются:
— хранение, копирование и экспрессия генетической информации;
— внутриклеточный транспорт;
— импорт питательных субстратов;
— конструктивный метаболизм, т. е. совокупность процессов ассимиляции субстратов и диссимиляции продуктов жизнедеятельности;
— биосинтез и таргетинг материала для образования клеточных структур;
— накопление запасных веществ и их расходование;
— экспорт экзометаболитов и отходов метаболизма.
Однако мембраны вакуома не способны ассимилировать энергию путем создания протонного градиента. Эту роль в ядерных клетках выполняют кристы митохондрий и тилакоиды пластид.
Ядерная оболочка изолирует сайты процессинга генетического материала от сайтов его экспрессии. Напомним, что репликация и транскрипция (вместе с посттранскрипционным процессингом, который включает в себя сплайсинг, кэппирование мРНК и т. д.) происходят в нуклеоплазме, а трансляция осуществляется в цитоплазме.
В отличие от эукариотов, у прокариотов отсутствует пространственно-временная изоляция транскрипции и трансляции, и их рибосомы присоединяются ко входному 5'-концу формирующейся молекулы мРНК, не дожидаясь финала образования транскрипта. Это происходит потому, что у прокариотов суммарная скорость инициации, транскрипции и элонгации выше, чем скорость считывания матрицы ДНК с помощью РНК-полимеразного комплекса.
Субъединицы эукариотной 80S рибосомы экспортируются через ядерные поры в цитоплазму, и только там образуется способный к трансляции комплекс рибосома- мРНК. Однако трансляция может происходить и внутри ядра — в 2001 г. Иборра (F. J.Iborra) с помощью иммуноцитохимических методов показал, что ядра млекопитающих в небольшом количестве используют аминоацил-тРНК. Смысл внутриядерной трансляции может заключаться в тестировании мРНК на наличие нонсенс- кодонов, чтобы обеспечить их «нонсенс-обусловленное» устранение (англ. nonsense- mediated decay, NMD).
5.2. Эволюционное происхождение эукариотов
Особое место в теоретической биологии занимает реконструкция сценария происхождения эукариотной клетки. Трудность в том, что вопрос о происхождении ядра стоит отдельно от вопроса о происхождении эукариотной клетки как отдельного эволюционного ствола.
В настоящее время рассматриваются две гипотезы о происхождении эукариотов.
В середине 1980-х годов господствовала идея о независимом происхождении эукариотов, бактерий и архей от гипотетического общего предка. Однако дендрограммы, отражающие результат секвенирования консервативных белков (факторов элонгации EF-G и EF-Tu, протон-транслоцирующих FoFi-АТФаз и V-АТФаз, молекулярного шаперона Hsp70, глутаматдегидрогеназы и т. д.), не совпадают с дендрограммами, построенными на основе сравнения рРНК. Поэтому единого мнения о том, как выглядит основание эволюционного древа, до сих пор не существует.
По наиболее известному сценарию, бактерии дивергировали с общим предком архей и эукариотов (см. рис. 2). Карл Воз, Ивабе (N. Iwabe), Гогартен (J. Р. Gogarten) и другие крупные авторитеты в области эволюционной цитологии считают, что предки эукариотов, или «протоэукариоты» имели такое же строение, как археи. Позднее они
приобрели вакуом, цитоскелет и, наконец, ядро. В этом случае гомология эукариотных и прокариотных факторов элонгации, протонных АТФаз и других важнейших белков объясняется «горизонтальным» переносом генов. Химерная природа ядерного генома может быть следствием обмена генетической информацией, который происходил либо между свободноживущими представителями трех доменов, либо после того, как бактерии стали облигатными эндосимбионтами и превратились в митохондрии и пластиды.
Альтернативный сценарий предложил в 1989 г. Вернер Циллиг. В данном случае химерная природа ядерного генома объясняется симбиотическим слиянием двух прокариотных линий. В начале 1990-х годов Джеймс Лейк, Хориике (Т. Horiike) и Гупта (R. Gupta) предположили, что ядро произошло от археотной клетки, которая путем фагоцитоза попала внутрь бактерии. Еще один сторонник археотного происхождения ядра, американский молекулярный биолог Соджин (М. Sogin) считает, что предшественником эукариотной клетки была не бактерия, а гипотетическая протоклетка. Она представляла собой автореплицирующуюся мембранную везикулу с примитивным РНК-овым геномом, копирование которого осуществлялось при помощи РНК-репликазы.
Сохранились ли в современной биосфере прямые потомки первичных ядерных организмов? Есть предположение, что это могут быть анаэробные протисты из группы Archezoa, у которых нет митохондрий, гидрогеносом, пластид, а также диктиосом и пероксисом (см. гл. 6).
Существует и противоположная точка зрения, согласно которой амитохондриальные протисты не являются прямыми потомками архаичных эукариотов (у их предков были митохондрии, но они потеряли их в результате вторичной адаптации к анаэробной среде обитания). В качестве аргумента приводится то, что в ядерном геноме амитохондриальных протистов обнаружены гомологи бактериальных генов. Однако это можно с равным успехом объяснить «горизонтальным» переносом генов между свободноживущими организмами на начальном этапе биологической эволюции.
5.3. Протоклетка
Отдельный тип монады — это гипотетическая протоклетка. Она находится в основании родословного древа и служит общим предком всех трех глобальных доменов. Воз назвал ее «прогенотом» (англ. progenitor — прародитель).
Реконструкция свойств прогенота, которого в последнее время называют «последним универсальным общим предком» (англ. Last Universal Common Ancestor, LUCA) — это отдельная проблема теоретической биологии. Перед исследователями стоит задача локализовать LUCA на глобальной дендрограмме, а также дать ретроспективное описание его биологических свойств.
Несколько крупных биологов, среди которых в первую очередь нужно назвать де Дюва, склоняются к мысли о том, что мир протоклеток в свое время пришел на смену миру эволюционирующих молекулярных существ. В 1986 г. Гилберт (W. Gilbert) назвал его «миром РНК» (англ. RNA world). Как полагают, в ту эпоху еще не существовали белки, и РНК одновременно играла роль носителя генетической информации и биокатализатора.
По мнению Отто Кандлера, клеточная жизнь первоначально была колониальной и недифференцированной. На заре эволюции «горизонтальный» перенос генов мог
быть строго закономерным, а не эпизодическим, как теперь, явлением. Современная биота сохраняет свое разнообразие благодаря трофической взаимозависимости индивидов при относительной стабильности геномов из-за существования генетических барьеров. В отличие от этого, биологической стратегией мира протоклеток был свободный обмен быстро мутирующими генами при использовании трофических резервов, накопленных абиотическим путем. Скорее всего, LUCA представлял собой не отдельный вид, а поливариантный продукт «горизонтального» переноса генов между разными формами жизни. По мнению канадского молекулярного биолога Уильяма Дулитла (W. F. Doolittle), мы никогда не сможем узнать, как выглядит основание древа жизни, поскольку скорость «горизонтального» переноса генов могла быть выше скорости дивергенции.
В отличие от Отто Кандлера и Уильяма Дулитла, а также американского молекулярного генетика Рассела Дулитла (R. W. Doolittle), большинство авторов считает, что хотя LUCA был достаточно примитивным, он имел все атрибуты клетки, как то:
— геном, представленный ДНК;
— набор ферментов и других белковых факторов, необходимых для биосинтеза ДНК и РНК;
— рибосомы, т. е. рибонуклеопротеиновые системы процессинга РНК-транскриптов;
— системы посттрансляционного процессинга белка, в том числе молекулярные шапероны;
— аппараты рекомбинации, генетической регуляции и репарации;
— механизмы контроля клеточного деления;
— системы регенерации АТФ и мембранную электрон-протонофорную цепь.
При попытке реконструировать фенотип LUCA неизбежно встает вопрос о характере адаптации протоклеток к физико-химическим условиям, которые господствовали на Земле 3,5 млрд лет назад. Согласно геологическим данным, тело нашей планеты сконденсировалось вокруг ее металлического ядра 4,5 млрд лет тому назад, а первобытный Океан сформировался 4,2 млрд лет тому назад. В течение следующих 500 млн лет земная кора несколько раз подвергалась «последней сильной метеоритной бомбардировке» (англ. Late Heavy Meteorite Bombardment, LHB), вследствие чего температура водоемов доходила до точки кипения, что способствовало отбору гипертермофильной биоты, и поэтому LUCA был термофилом.
Идут споры том, какое место занимает LUCA на глобальной дендрограмме. По консенсусной версии, корень клеточной жизни расположен где-то между стволом бактерий и другим стволом, который в дальнейшем разветвился на стволы архей и эукариотов (см. рис. 2). Скорее всего, LUCA был похож на бактерию, и можно реконструировать его фенотип, анализируя полностью секвенированные бактериальные геномы.