ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 11. Взаимодействие клеток при гуморальном иммунном ответе
ДЕЙСТВИЕ ЦИТОКИНОВ НА В- И Т-КЛЕТКИ
Главная роль в активации Т-клеток принадлежит ИЛ-2
Пролиферация лимфоцитов — это сложный, опосредуемый многими факторами процесс. Так, стимуляция Т-клеток АПК еще не означает, что лимфоциты сразу же будут пролиферировать. Эффективное взаимодействие с участием ТкР ведет к продукции α-цепи (р55) рецептора для фактора роста Т-клеток, ИЛ-2. α-Цепь и уже имеющиеся β (р75)- и γ-цепи ассоциируют с образованием высокоаффинного рецептора для ИЛ-2 (рис. 11.16). Активация ТкР у всех Т-клеток вызывает продукцию цитокинов. Большинство Т-клеток CD4+и некоторые Т-клетки CD8+секретируют ИЛ-2 непродолжительное время, 1-2 сут. В течение этого срока взаимодействие ИЛ-2 с высокоаффинным рецептором для ИЛ-2 приводит к росту и активации Т-клеток. После стимуляции ТкР клетки экспрессируют высокоаффинный рецептор только в течение примерно одной недели: благодаря такому ограничению Т-клетки не размножаются беспредельно, так как отсутствие положительных сигналов способствует индукции апоптотической гибели лишних клеток. Схема всего процесса активации Т-клеток представлена на рис. 11.17.
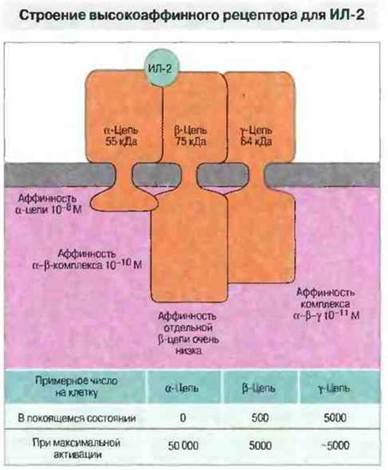
Рис. 11.16. Высокоаффинный рецептор для ИЛ-2 состоит из 3 цепей, каждая из которых в отдельности лишь слабо связывает ИЛ-2. Покоящиеся Т-клетки не экспрессируют α-цепь, но после активации они могут экспрессировать до 50 000 α-цепей на клетку (максимальное число). Некоторые из этих цепей соединяются с β-цепью, образуя высокоаффинный рецептор для ИЛ-2.
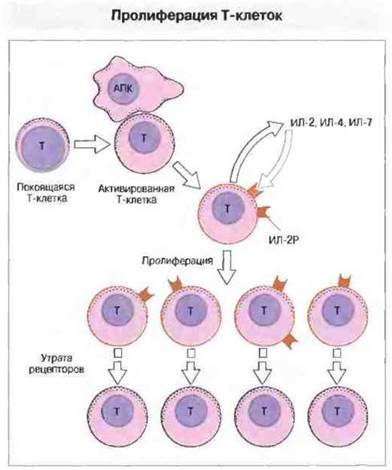
Рис. 11.17. Покоящиеся клетки не продуцируют цитокинов - факторов роста Т-клеток (ИЛ-2, ИЛ-4 или ИЛ-7) и экспрессируют лишь небольшое число рецепторов для ИЛ-4 и ИЛ-7, но не для ИЛ-2. На их поверхности присутствует только цепь р75 рецептора ИЛ-2, обладающая низкой аффинностью. Активация ведет к появлению цепи р55, которая вместе с р75 образует высокоаффинный рецептор ИЛ-2.
Активация индуцирует синтез мРНК и белков ИЛ-2 и ИЛ-4. Секреция этих цитокинов и их взаимодействие с рецепторами вызывают пролиферацию. Цитокины действуют двумя способами: «аутокринным» (влияя на те клетки, которые их продуцируют) и «паракринным» (стимулируя соседние клетки). В отсутствие антигенной стимуляции число рецепторов для ИЛ-2 уменьшается и пролиферативная фаза иммунного ответа заканчивается.
ИЛ-2 синтезируют главным образом Т-клетки CD4+, однако его продуцентами служат также СD8+-клетки и большие гранулярные лимфоциты (БГЛ). Действие ИЛ-2 ограничено определенными видами клеток, в основном Т-клетками всех типов, для которых данный цитокин служит наиболее сильным фактором роста и активатором (рис. 11.18). Он влияет также на БГЛ и В-клетки, индуцируя их рост и дифференцировку, активирует макрофаги и, возможно, олигодендроциты. В виде препарата ИЛ-2 используют для экспериментальной терапии опухолей, в частности почечно-клеточного рака. Лечебное действие этого цитокина может быть связано с активацией различных клеток, обладающих противоопухолевым цитотоксическим действием, например лимфокин-активированных клеток-киллеров (ЛАК-клеток).
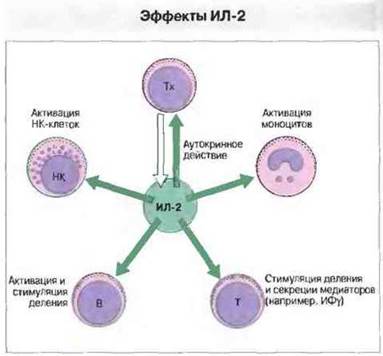
Рис. 11.18. ИЛ-2 образуют Тх-клетки. Функции этого цитокина включают стимуляцию деления Т-клеток, высвобождения медиаторов, таких как ИФγ, индукцию роста В-клеток, а также активацию моноцитов и НК-клеток для усиления иммунного ответа. Аутологичные предшественники НК-клеток больных почечно-клеточным раком могут быть активированы in vitro высокими дозами ИЛ-2 (1000 ИЕ/мл) для получения линий так называемых лимфокин-активированных клеток-киллеров (ЛАК-клетки), которые используются для экспериментальной терапии рака.
Менее сильным фактором роста Т-клеток служит ИЛ-4, который также относится к числу индуцибельных цитокинов. Экспрессия поверхностного рецептора для ИЛ-4 повышается в результате активации ТкР. На Т-клетки действует также ИЛ-7, хотя первоначально он был описан как фактор роста пре-В-клеток, образуемый стромой костного мозга. Продуцируемый элементами стромы тимуса ИЛ-7 действует на тимоциты, представляя собой фактор роста и активации Т- клеток в дополнение к свойству активировать макрофаги. Возможно, описанные три фактора роста, а также менее изученные цитокины, в частности ИЛ-9, ИЛ-12 и ИЛ-15, осуществляю! тонкую регуляцию роста и активации Т-клеток в ходе иммунного ответа.
Цитокины ИЛ-1 и ИЛ-6 служат костимулирующими сигнальными молекулами при активации Т-клеток некоторыми АПК (рис. 11.19). Удивительно, что еще никому не удалось обнаружить продукцию этих цитокинов теми АПК, которые считаются наиболее эффективными презентирующими клетками при первичной Т-клеточной активации, — интердигитатными дендритными клетками. Не исключено действие в качестве костимулирующих и других цитокинов, включая ИЛ-12 (образуемый дендритными клетками) и ИЛ-15.
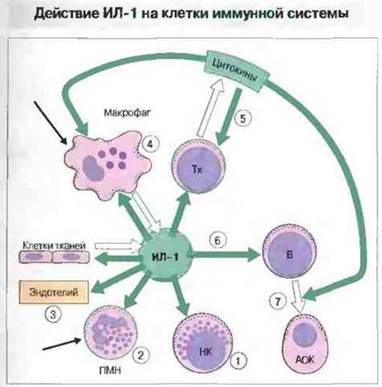
Рис. 11.19. ИЛ-1 продуцируют клетки многих типов в ответ на повреждение, инфекцию или антиген. Он влияет на разнообразные клетки и процессы, вызывая 1) возрастание цитотоксической активности НК-клеток; 2) повышение метаболической активности полиморфноядерных нуклеоцитов (ПМН), которые мигрируют в район продукции ИЛ-1 (за счет хемотаксиса) (черная стрелка); 3) индукцию синтеза молекул адгезии и прокоагулянтов в эндотелиальных клетках и увеличение проницаемости эндотелия; 4) повышение продукции простагландинов и цитокинов макрофагами и их цитоцидной и хемотаксической (черная стрелка) активности; 5) усиление пролиферации Тх-клеток, экспрессии рецепторов ИЛ-2 и продукции цитокинов и 6) пролиферацию В-клеток и их дифференцировку в АОК (оба эти процесса регулируются другими цитокинами (7)).
Цитокин ИЛ-3 стимулирует рост клеток- предшественников всех гемопоэтических линий (эритроцитов, гранулоцитов, макрофагов и, возможно, лимфоцитов). На этот цитокин реагирует также минорная популяция Т-клеток (Т-клетки CD4-CD8-, несущие αβ-ТкР).
Активацию и деление В-клеток индуцируют цитокины, образуемые Тх2-лимфоцитами
К специфическим цитокинам, продуцируемым Тх2-клетками, относятся ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10 и ИЛ-13. Цитокин ИЛ-4 (первоначально известный как фактор 1 В-клеточной активации или дифференцировки) действует на В-клетки, индуцируя их активацию и дифференцировку с преимущественной продукцией антител IgG I и IgE. Он влияет также на Т-клетки как фактор роста, способствуя дифференцировке Тх2-лимфоцитов и усиливая тем самым антителообразование. Одновременно он ингибирует секрецию макрофагами провоспалительных цитокинов, таких как ИЛ-1 и ФНОα. Избыток ИЛ-4, вызывающего синтез IgE, играет патогенетическую роль при аллергических заболеваниях.
ИЛ-5 функционирует в организме человека в основном как фактор роста и активации эозинофилов. У мыши он действует также на В-клетки, индуцируя их рост и дифференцировку. Этот цитокин обусловливает эозинофилию при паразитарных инвазиях.
Цитокин ИЛ-6, известный ранее как фактор В-клеточной дифференцировки или фактор стимуляции гепатоцитов, образуют многие клетки — Т-лимфоциты, макрофаги, В-клетки, фибробласты и эндотелиальные клетки. Он действует на самые разнообразные клетки, но особенно важна его функция как индуктора дифференцировки В-лимфоцитов в антителообразующие клетки (АОК). В печени ИЛ-6 стимулирует синтез белков острой фазы. Считается, что ИЛ-6 служит важным фактором роста множественной миеломы — злокачественного новообразования, возникшего из плазматических клеток.
Цитокины - регуляторы иммунных реакций
От цитокинов может также зависеть тип развивающегося иммунного ответа. Особенно важна при этом регуляторная функция ИЛ-10 и ИЛ-12. Цитокин ИЛ-10 (известный также как фактор, ингибирующий синтез цитокинов) подавляет синтез ИФγ. Кроме того, он ингибирует презентацию антигенов и синтез ИЛ-1, ИЛ-6 и ФНОα макрофагами, что благоприятствует развитию иммунного ответа по Тх2-типу и В-клеточной активации. Действие ИЛ-12 комплементарно эффектам ИЛ-10: он способствует развитию Тх1-ответа, сопровождающегося активацией макрофагов и нормальных киллерных клеток (НК-клеток).
Источником образования ИФγ служат активированные Т-лимфоциты и НК-клетки. Иммунная активация ведет к образованию ИФγ и повышению антигенпрезентирующей функции клеток многих типов, в том числе макрофагов, астроцитов, клеток микроглии и эндотелия, а также тимоцитов. Частично этот эффект опосредован усилением экспрессии генов МНС классов I и II и ТАР, которое влечет за собой дальнейшую активацию Т-клеток. Повышается также экспрессия костимулирующих молекул В7-1, В7-2 и ICAM-1. Таким образом, при многих иммунных реакциях ИФγ выполняет функцию положительного сигнала регуляции по механизму образной связи. Однако вместе с тем он ингибирует пролиферацию Тх2-клеток, как правило активирует макрофаги и, как это ни парадоксально, препятствует индукции синтеза МНС-молекул класса II В-клетками, вызываемой ИЛ-4. Отсюда следует, что ИФγ способствует развитию иммунного ответа Тх1-типа. Свойства ИФγ иллюстрирует рис. 11.20.
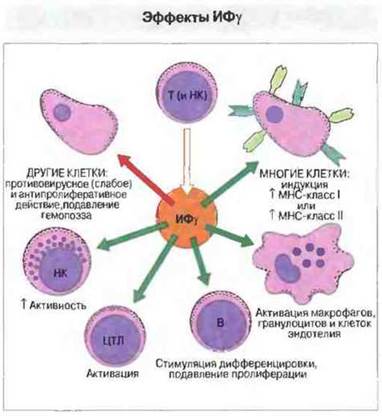
Рис. 11.20. Интерферон γ (ИФγ) оказывает многочисленные иммунорегуляторные эффекты. Его противовирусное и антипролиферативное действие менее выражено, чем у ИФα и ИФβ. Кроме того, он не столь эффективно стимулирует НК-клетки, как ИФα. Однако при этом ИФγ служит наиболее сильным активатором макрофагов и индуктором экспрессии молекул МНС класса II клетками тканей. Осуществляя эти и другие функции, ИФγ действует синергично с ФНОα и ФНОβ.
ИФα и ИФβ, подобно ИФγ, могут подавлять пролиферацию клеток иммунной системы. Они ингибируют, кроме того, эффект ИФγ, индуцирующий экспрессию МНС-антигенов класса II. Цитокин ИЛ-6, стимулируя дифференцировку клеток, в том числе некоторых лейкозных линий, тормозит их пролиферацию. Цитокины ИЛ-6 и ИЛ-11 способны ингибировать синтез макрофагами провоспалительных цитокинов, однако их действие слабее, чем у ИЛ-4 или ИЛ-10. Цитокин ИЛ-13 по своим свойствам подавлять функции макрофагов во многом сходен с ИЛ-4.
Трансформирующий фактор роста β (ТФРβ) представлен семейством из трех близкородственных молекул, которые стимулируют рост соединительной ткани и образование коллагена, но ингибируют практически все иммунные и гемопоэтические функции, особенно если присутствуют до активации клеток.