ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 1. Общие сведения об иммунной системе
ИММУННЫЙ ОТВЕТ
Итак, любой иммунный ответ имеет две основные фазы:
✵ распознавание антигена;
✵ реакции, направленные на его устранение.
В реакциях приобретенного иммунитета распознавание антигена осуществляют лимфоциты, избирательно пролиферирующие благодаря клональной селекции.
Клональная селекция - пролиферация клеток, связавших специфический антиген
Каждый лимфоцит (как В-, так и Т-популяции) генетически запрограммирован распознавать в основном только один антиген, но иммунная система в целом может специфически распознать многие тысячи разных антигенов. Поэтому лимфоциты, способные распознать тот или иной антиген, должны составлять лишь очень малую часть общей популяции. Как же в таком случае организм адекватно отвечает на инфекцию? Объяснение состоит в том, что антиген, связавшись с теми немногими клетками, которые способны его распознать, вызывает их быструю пролиферацию. В течение нескольких дней появляется достаточно клеток для адекватного иммунного ответа. Иными словами, сам антиген выбирает и способствует образованию специфических клонов клеток, связывающих этот антиген (рис. 1.13), — процессу, названному клональной селекцией и свойственному как В-, так и Т-клеткам.
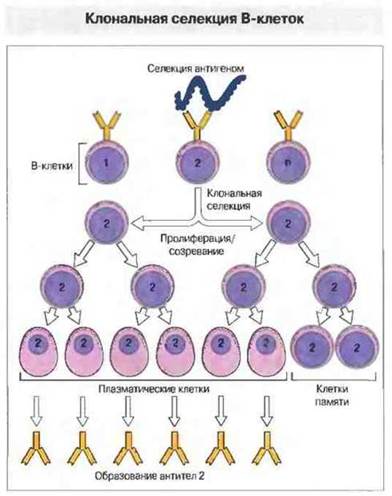
Рис. 1.13. Каждая антителообразующая клетка (В-клетка) запрограммирована синтезировать антитела только одной специфичности. Они расположены на ее поверхности в виде антигенсвязывающих рецепторов. Антиген связывается только с теми В-клетками, которые несут соответствующий поверхностный рецептор - в нашем примере В-клетка 2. Это взаимодействие стимулирует пролиферацию таких клеток и их созревание в клетки, образующие антитела, а также в долгоживущие клетки иммунологической памяти, все с той же исходной специфичностью связывания антигена.
Кажется непостижимым, каким образом иммунная система может «предугадать» репертуар специфичностей антител, которые потребуются в течение будущей жизни индивида. На самом деле все обстоит иначе. Просто иммунная система производит антитела, способные распознать огромное разнообразие антигенов, еще до встречи с ними. Многие из этих антител никогда не будут востребованы для защиты данного индивида от инфекции. Однако бесчисленное множество патогенных микроорганизмов и их способность к изменению своего антигенного состава в результате мутаций делает наличие всех этих антител необходимым — на случай, когда они могут понадобиться.
Лимфоциты, активированные связыванием антигена, вступают в цикл клеточного деления. Они экспрессируют новые рецепторы, позволяющие им реагировать на выделяемые другими клетками цитокины, которые служат сигналами к пролиферации. Лимфоциты могут также сами начать выделение цитокинов. Обычно они проходят ряд циклов деления, прежде чем дифференцируются в зрелые клетки, снова под действием цитокинов. Например, пролиферирующие В-клетки в итоге созревают в образующие антитела плазматические клетки. Даже после устранения инфекции сохраняется некоторая часть новообразованных лимфоцитов, способных вновь активироваться, если антиген встретится им повторно. Их называют клетками памяти, гак как они хранят иммунологическую память относительно отдельных антигенов. Существованием клеток памяти и обусловлен долгосрочный иммунитет к тому или иному возбудителю.
Для устранения патогенных микроорганизмов существуют различные эффекторные механизмы иммунного ответа
Иммунная система располагает множеством механизмов для разрушения патогенных микробов, и каждый из них соответствует данному типу инфекции и конкретной стадии жизненного цикла возбудителя. Эти механизмы защиты часто называют эффекторными системами.
Нейтрализация. При действии одной из самых простых эффекторных систем антителам достаточно только связаться с определенным возбудителем, чтобы оказать ему противодействие. Например, антитела к наружным белкам капсида некоторых риновирусов (вызывающих простудные заболевания) могут воспрепятствовать связыванию вирусных частиц с клетками организма и их инфицированию.
Фагоцитоз. Гораздо чаше антитела реализуют свой эффект, активируя комплемент или действуя в качестве опсонинов, усиливающих поглощение микробов фагоцитами. Связавшись с опсонизированным микробом, фагоцитарная клетка поглощает его, окружая выступающими псевдоподиями. Псевдоподии сливаются, и микроб оказывается заключенным (эндоцитированным, интернализованным) в фагосому (рис. 1.14). Перерабатывают фагоциты поглощенный материал по-разному. Макрофаги, например, восстанавливают молекулярный кислород с образованием бактерицидных реакционноспособных метаболитов кислорода, которые секретируются в фагосому. Нейтрофилы содержат лактоферрин, который хелатирует железо, лишая некоторые бактерии этого необходимого элемента питания. Наконец, с фагосомой сливаются гранулы и лизосомы, наполняя возникшую фаголизосому ферментами, разрушающими ее содержимое (рис. 1.15). Более полно механизмы фагоцитоза описаны в гл. 10 и 17.
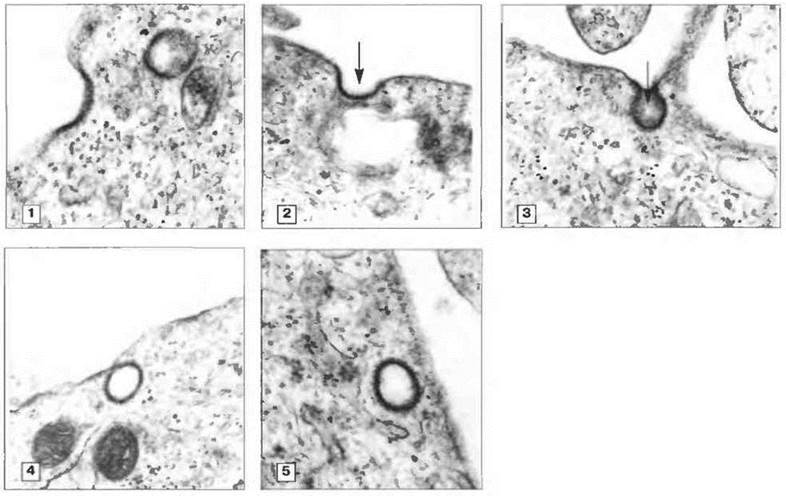
Рис. 1.14. Электронно-микроскопическое изучение фагоцитоза. На микрофотографиях представлены последовательные стадии инвагинации мембраны и происходящей в результате этого интернализации материала. Фагоцит, связавшись с опсонизированной частицей, постепенно поглощает ее, выпуская псевдоподии (1-3). Затем псевдоподии сливаются (4), и микробная клетка оказывается заключенной в фагосому (5). (Фото любезно предоставлены д-ром A. Stevens и проф. J. Lowe.)
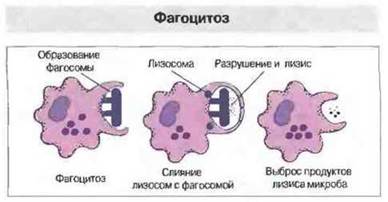
Рис. 1.15. Фагоциты поступают в очаг воспаления благодаря хемотаксису. Затем их поверхностные неспецифические рецепторы связываются с микробами, либо, если микробная поверхность опсонизирована фрагментом третьего компонента комплемента (С3b) и/или антителами, связывание происходит с участием фагоцитарных рецепторов для С3b и/или Fc (см. рис. 1.10). Когда в результате связывания фагоцит активируется, он окружает инфекционный агент псевдоподиями, заключая в фагосому; при этом происходит образование бактерицидных метаболитов кислорода. Как только микроб поступит внутрь клетки, лизосомы сливаются с фагосомой, образуя фаголизосому, в которой инфекционный агент уничтожается. Остатки микроба могут быть выделены клеткой наружу.
Цитотоксические реакции и апоптоз. Цитотоксические реакции — это эффскгорные иммунные механизмы, направленные против целых клеток, обычно против тех, которые слишком крупны для фагоцитоза. Такая клетка-мишень распознается либо специфичными антителами, взаимодействующими с компонентами ее поверхности, либо Т-клетками посредством антигенспецифичных ТкР. В отличие от фагоцитоза, при котором содержимое лизосом изливается в фагосому, в цитотоксической реакции атакующая клетка направляет содержимое своих гранул наружу, к клетке-мишени. Гранулы цитотоксических Т-клеток содержат соединения, называемые перфоринами, которые способны создавать каналы в наружной мембране клеток-мишеней. (Подобно этому, антитела, связавшись с поверхностью клегки-мишени, могут привлечь комплемент для перфорирования ее цитоплазматической мембраны.) Некоторые цитотоксические клетки способны также своим сигналом включать программу саморазрушения клетки-мишени — процесс апоптоза.