Эволюция вирусов - Жданов В. М. 1990
Специальная часть
Парамиксовирусы
Парамиксовирусы вместе с ортомиксо-, рабдо-, бунья- и аренавирусами образуют четко ограниченную группу вирусов с негативно-полярным геномом. Их РНК неинфекционная и не кодирует непосредственно синтез белков. Последнему предшествует синтез комплементарной нити, которая выполняет функцию мРНК, причем независимо от того, является геном непрерывистым или прерывным, каждый ген транскрибируется отдельно, имея свои рамку считывания и регуляторную область. Естественно, что синтез комплементарной нити (или нитей), выполняющей функции мРНК, обеспечивается вирусспецифической полимеразой, находящейся в вирионах. Эта же полимераза (или полимеразный комплекс) синтезирует полную нить (или нити) комплементарной РНК, которая служит матрицей для синтеза дочерних РНК.
Таковы общие признаки этих 5 групп вирусов, отличающие их от вирусов любой другой группы. Между ними имеются и различия. Парамиксо- и рабдовирусы обладают непрерывным геномом, у аренавирусов он состоит из двух фрагментов, у буньявирусов — из 3, а у ортомиксовирусов — из 7—8. Различна и общая их величина: до 4,6x106у рабдовирусов, до 4,8х106у аренавирусов, 5x106у ортомиксовирусов, 5х106—7x106у парамиксовирусов и буньявирусов. Размеры вирионов при сферической форме 90—100 нм (буньявирусы), 80—120 нм (ортомиксовирусы), 110—130 (арена-вирусы), 140—150 (парамиксовирусы); пуле- или палочковидные вирионы рабдовирусов имеют размер 50—90х130х380 нм. Нуклеокапсид (рибонуклеопротеид) упакован по спиральному типу симметрии. Круг поражаемых «хозяев» — теплокровные животные (парамиксо-, ортомиксо- и аренавирусы), теплокровные животные или кровососущие переносчики (бунья- и рабдовирусы). Большая группа последних вирусов поражает растения и передается сосущими насекомыми. Описаны также морфологически сходные вирусы, обнаруженные у растений, у них же выявлены морфологически сходные структуры без оболочек (см. главу 14).
Сходство основных характеристик 5 групп вирусов с негативно-полярным геномом позволяет думать о едином или близком их происхождении, а круг «хозяев» — высшие животные и растения, а также насекомые-переносчики (кровососы или сокососы) — все это дает основание предполагать, что появились эти вирусы не ранее 200—300 млн лет назад, и, может быть, значительно позже. В то же время по крайней мере одна из рассматриваемых групп успела заселить обширные экологические ниши (высшие животные, высшие растения).
О происхождении вирусов с негативно-полярным геномом можно высказать несколько предположений. Возможно, они возникли из клеточных структур и далее дивергировали вплоть до формирования ныне существующих 5 групп. Возможно, у них не было единого предка, а сходные структуры возникли повторно и затем самостоятельно эволюционировали. Но нам представляется более вероятным предположение о происхождении их от вирусов с двухнитевой РНК — первой «защитой» автономного РНК-генома от действия клеточных нуклеаз. Такими структурами являются дрожжевые киллеры. Дальнейшим этапом явились вирусы грибов, имеющие геном в виде двунитевой РНК и уже собственную РНК- полимеразу. Заметим, что эти вирусы могли появиться очень давно, когда возникли примитивные формы эукариотов — грибы. Реовирусы могли стать одной из магистральных ветвей эволюции примитивных вирусов с двунитевой РНК. Их обилие, а также заселение обширной экологической ниши (высшие животные, высшие растения, переносчики — членистоногие) свидетельствуют о реализации такого эволюционного направления. Но другим направлением мог стать возврат к геному, состоящему из однонитевой РНК, так как в вирионе такой геном хорошо защищен от действия клеточных нуклеаз, а репликативными формами по-прежнему является двунитевая РНК. Естественно, при наличии вирионной 'полимеразы геномом должна была стать минус-нить, что и было реализовано в данном эволюционном направлении. Отметим, между прочим, что ни один из вирусов с однонитевой РНК и вирионной полимеразой не имеет положительно-полярного генома, не считая, конечно, ретровирусов.
В соответствии с этим предположением существующие 5 групп вирусов с негативно-полярной геномной РНК вряд ли имели единого предка, скорее их было несколько, но тем не менее они были эволюционно близкими. В пользу такого предположения свидетельствуют ограниченные колебания молекулярной массы генома (от 4х106до 7х106). Среди них нет «карликов» вроде мелких РНК-содержащих фагов (масса генома около 106). Размер генома не выходит за пределы 7х106—8х106 у всех РНК-содержащих вирусов, даже у реовирусов, для которых величина 15х106—16х106 на самом деле означает «половинный» размер, так как генетическая информация закодирована только на одной нити РНК.
Однако на самом деле эволюционный путь, который проделали эти вирусы (в частности парамиксовирусы), мог быть гораздо сложнее. Об этом свидетельствуют данные по получению моноклональных антител против белка F вируса кори. Из 11 таких антител 3 реагировали с белком (79 000) клеток HeLa, синтез которого был индуцирован другими парамиксовирусами, тепловым шоком, канамицином и др. Этот белок называют стрессовым [Sheshbezadaran Н., Norrby Е., 1984]. Цитируемые авторы рассматривают данный феномен как возможную основу аутоиммунных реакций, мы же хотим поставить здесь вопрос — идет ли речь об эволюционной общности перекрестнореагирующих белков или имеет место молекулярная конвергенция.
После этих соображений о происхождении РНК-содержащих вирусов с негативно-полярным геномом рассмотрим эволюцию парамиксовирусов; предварительно напомним некоторые необходимые сведения о них [Закстельская Л. Я-, Зайдес В. М., 1982; Matthews R., 1982].
Как уже указывалось, геном парамиксовирусов представляет собой однонитевую негативно-полярную РНК с молекулярной массой 5х106—7х106 и коэффициентом седиментации 45—575. Некоторые вирионы (до 30%) содержат плюс-нити, которые надо рассматривать как результат небалансированных синтезов. На 5'-конце молекулы имеется поли (А)-последовательность. Вирионная РНК неинфекционна, но рибонуклеопротеид обладает инфекционными свойствами. Матричные РНК гетерогенны и седиментируют при 18—35 5 [Kingsbury D. et al., 1978].
Ген Р вируса кори содержит 1657 нуклеотидов и может кодировать синтез белка, состоящего из 507 аминокислотных остатков. На самом же деле он кодирует синтез двух белков, причем второй из них (С), содержащий 186 аминокислотных остатков, синтезируется с совпадающей рамкой считывания [Bellini W. et al., 1985]. Этот феномен имеет место и у других парамиксовирусов, в частности вируса Сендай (рис. 22). Возможно, что белок С играет определенную роль в предполагаемой ядерной фазе репликации вируса кори. Внутренние белки NP, Р и М парамиксовирусов (вируса Сендай) расположены следующим образом: белки NP — вдоль всей нити РНК, белок Р формирует дискретные кластеры в разных местах нуклеокапcида, белок М в цитоплазме зараженных клеток образует скопление вдоль нуклеокап- сида, а в вирионах окружает нуклеокапсид [Portner А., Murti К., 1986].
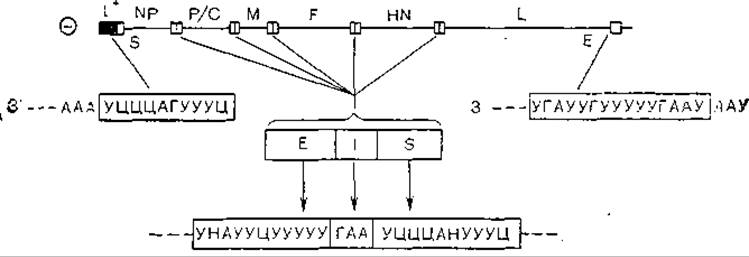
Рис. 22. Структура генома вируса Сендай. Указано положение плюснитевой лидерной последовательности (l+); Е, І и 5 обозначают концевые соответственно интергенные и стартовые последовательности мРНК..
Из 5—7 вирионных белков с молекулярной массой от 35 000 до 200 000 отметим нуклеопротеид NP (60 000), РНК-зависимую РНК-полимеразу (белки Р и L с молекулярной массой соответственно 75 000 и 160 000), белки внутренней мембраны М (40 000—43 000), гемагглютинин-нейраминидазу HN (67 000—76 000), фактор гемолиза и слияния клеток F (55 000—60 000). Современные данные указывают на наличие в сердцевинах вирионов 5 белков: NP0 (68 000), NP1 (58 000), NP2 (52 000), которые ассоциируются с 505 РНК, L (240 000) —полимераза и Р (84 000) —фосфопротеид. Четыре белка находятся в оболочках: F1 + F2 (53 000+12 000), HN (72 000) и М (40 000) [Jambou R. et al., 1985].
У респираторно-синцитиального вируса после промотора гены расположены следующим образом: З'-IC—IB—N—Р—М—IA—G—F—22 000—L-5'. Ген N кодирует синтез нуклеопротеида, G и F — поверхностных гликопротеидов, М — мембранного белка, Р и L — полимеразного комплекса. Гены разделены последовательностями длиной 1—52 нуклеотида, функции остальных генов не вполне ясны. Для сравнения приведем расположение генов у рабдовирусов и двух родов парамиксовирусов. У рабдовируса (вирус везикулярного стоматита): N—NS—М—G—L; у парамиксовирусов (вирус Сендай и вирус парагриппа человека типа 3): NP—Р + С—М— —F—HN—L; ОВ5: NP—P+Y—M—F—SH—HN—L.
В общем, несмотря на различия орто- и парамиксовируcов (в частности, фрагментарный геном у первых и непрерывный у вторых), спектр белков у них на редкость сходный, включая гликопротеиды внешней оболочки, которые расщепляются на субъединицы и затем соединяются дисульфидными связями, принимая окончательную конформацию. Похожи и морфология вирионов, и в основных чертах цикл репродукции.
Парамикоовирусы сравнительно немногочисленны и поражают исключительно высших животных — млекопитающих и птиц. Среди них три рода — собственно парамиксовирусы (12 видов), морбилливирусы (4 вида), пневмовирусы (3 вида). Эволюцию можно представить как рассеивание по разным видам «хозяев», причем одни из них вызывают острые инфекции, другие — хронические и персистирующие инфекции.
Антигенные взаимоотношения между парамиксовирусами птиц и свиней представлены на рис. 23 [Lipkind М. et al., 1986].
Объединяя происхождением рабдо- и парамиксовирусы, А. Д. Альтштейн и Н. В. Каверин (1980) считают, что рабдовирусы произошли от вирусов растений и лишь позже их круг «хозяев» расширился, охватив сначала насекомых, а затем позвоночных. Приобретение способности размножаться в клетках респираторных органов привело к появлению парамиксовирусов.
Имеются основания считать орто- и парамиксовирусы эволюционно родственными. Это видно из сравнительных данных, указывающих на гомологию некоторых генов вируса Сендай и вируса гриппа типа A [Giorgi С. et al., 1983] (степень гомологии отмечена разным размером букв).
Гены вируса Сендай:
NP; agacaggattTTAGGGTcAAAGtATc CACccTgA GGagCA GGttcCAg ACCC.
P/C; TAAGAAAAACTTAGGGTgAAAGttcATcCAC TgA tcGGctCA GG CAaggccacACCC,.
M; TAAGAAAAACTTAGGGTgAAA GaaÄTttCACc TaAcaGGcgCAatGG CAgatatctat.
Ген NP вируса гриппа:
AGcAAAA gcAGGGT atata ATctCAC TgAgt GG CA tcCA tatc.
В то же время некоторые гены парамиксовирусов несут «печать» клеточного происхождения, что выражается в своеобразной антигенной мимикрии, выявленной у нуклеопротеидов вируса кори и респираторно-синцитиального вируса [Norrby Е. et al., 1986].
Основными критериями для выделения родов являются наличие нейраминидазной активности (парамиксовирусы), размеры нуклеокапсида (морбилливирусы) и другие структурные особенности (пневмовирусы). Мюрбилливирусы, кроме того, образуют группу серологически родственных вирусов. Уместно также отметить, что некоторые эти вирусы полипатогенны. Например, вирус парагриппа типа 3 поражает человека, овец, коров; вирус парагриппа типа 5 — обезьян, собак и птиц; вирус парагриппа типа 1 —человека и мышей. (Речь идет не о лабораторных экспериментах, а естественных эпидемических и эпизоотических процессах.) Кроме того, некоторые парамиксовирусы, поражающие животных, могут вызвать и заболевания людей. Так, вирус болезни Ньюкасла, поражающий кур, может вызвать у человека скоропроходящий конъюнктивит.
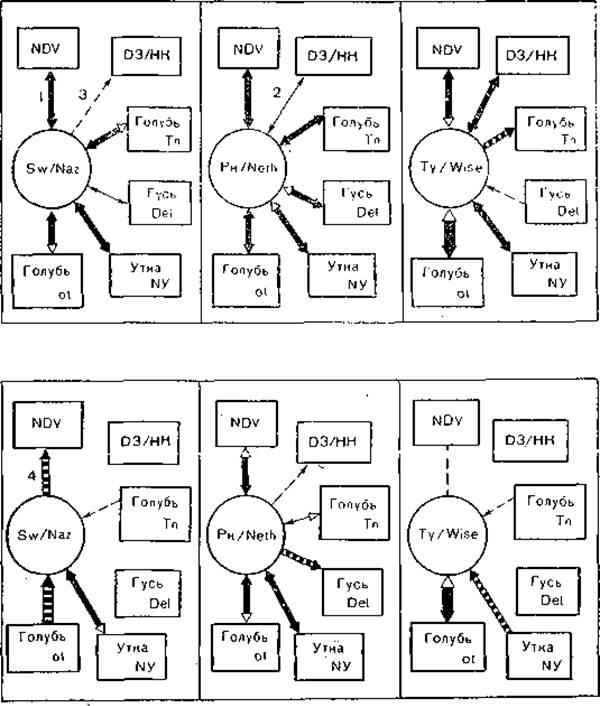
Рис. 23. Антигенная взаимосвязь парамиксовирусов. Стрелки направлены от вируса, чья антисыворотка ингибирует другой вирус, по направлению к вирусу, чья антисыворотка не ингибирует.
I — интериммунные сыворотки; II — сыворотки переболевших животных; 1 — значительное родство; 2 — умеренное родство; 3 — слабое родство; 4 — односторонняя связь.
Нелишне также напомнить, что среди круга поражаемых животных преобладают одомашненные человеком (рогатый скот, собаки, куры), а также заселяющие жилища человека мыши. Поэтому эту группу вирусов следует считать сравнительно поздним «образованием» — не более 10 000 лет назад, а вирусы, поражающие человека, появились много позже. О дериватном происхождении парамиксовирусов человека свидетельствует выраженный консерватизм генов HN вирусов коров и человека [Coelingh К. et al., 1986].
Остановимся на эволюции вируса кори, который относится к роду морбилливйрусов семейства парамиксовирусов. Попутно затронем вопрос о латентных вирусных инфекциях. При этом речь будет идти не о возможном филогенезе возбудителей этих инфекций, а об условиях их возникновения и эволюции на разных этапах развития человеческого общества.
Характерными особенностями всех инфекций, входящих в эту группу, являются легкость заражения, высокая восприимчивость к ним человека, острое течение, относительно короткий заразный период (от нескольких дней до нескольких недель), отсутствие носительства и стойкий постинфекционный иммунитет. Все это свидетельствует о невозможности существования данных инфекций при первобытнообщинном строе, когда люди жили разрозненно, мало связанными друг с другом и с другими племенами. Появление такой инфекции привело бы к поголовной заболеваемости племени в течение короткого срока, вслед за чем возбудитель неизбежно должен был погибнуть, даже если бы он отличался большой стойкостью (чего нет среди возбудителей этой группы болезней). Не только кочевой образ жизни, но и переход к оседлости при относительной, изолированности племен и малой численности населения еще не обеспечивали необходимых условий для сохранения паразитических видов этой группы.
Правильность этих выводов лучше всего иллюстрируется эпидемиологией кори. Корь — одна из самых широко распространенных детских болезней, и до проведения массовых иммунизаций коревой вакциной каждый человек в детстве переносит эту инфекцию. Высокая заразность больного корью и воздушно-капельный путь передачи инфекции приводят к тому, что в современном обществе человек в течение жизни многократно заражается этой инфекцией начиная с раннего детства. Вследствие абсолютной восприимчивости после первого же заражения, наступающего в детстве, ребенок заболевает. Перенесенная инфекция формирует стойкий иммунитет на всю жизнь, вследствие чего повторные заболевания не Наступают. Это позволяет сделать вывод: чем более скученно живет население и чем более выражено взаимное общение, тем в более раннем возрасте наступает заболевание. В городах с большой платностью населения дети обычно переносят корь їв возрасте до 3—4 лет. В сельских местностях, особенно если они оторваны от железных дорог и других путей сообщения, основная масса заболеваний корью передвигается на более поздние возрастные группы детей. Наконец, в местностях, изолированных от остального мира, корь не является детской болезнью, и возникающие при заносе кори эпидемии охватывают как детское, так и взрослое население.
Классическим примером являются эпидемии кори на Фарерских островах в XIX в. Первая эпидемия возникла после 65-летнего перерыва, на протяжении которого на островах не было ни одного случая кори. В марте 1846 г. на один из островов прибыл человек, перед отъездом имевший контакт с больным корью. Через несколько дней после прибытия он заболел корью, вслед за чем началась эпидемия кори, поразившая все население островов. Эпидемия прекратилась после того, как в окружении больных не осталось лиц, восприимчивых к этой инфекции. После этого на островах в течение 16 лет кори не было. Новая эпидемия вспыхнула в 1882 г. после заноса кори больным, прибывшим на остров. На этот раз инфекцией было охвачено население в возрасте до 16 лет, так как остальное население переболело во время предыдущей эпидемии. Еще через 13 лет, в 1875 г., вновь была «занесена» корь из метрополии, и на этот раз эпидемия охватила детей только в возрасте до 13 лет. Таким образом, по мере усиления связей с метрополией учащался «занос» кори на острова и эта инфекция становилась только детской. Однако корь не смогла удержаться на островах с населением 7000 человек, и каждая новая эпидемия возникала вследствие заноса возбудителя.
Приведенный пример показывает, что инфекция типа кори может существовать лишь на высокой стадии, развития человеческого общества, когда образовались крупные государства с обширными территориями, большими массами населения и развитыми путями сообщений. Высокая плотность населения и развитые пути сообщения обеспечивают распространение инфекции на территории, а обширность территории и значительное число населения приводят к тому, что, пока эпидемия успевает распространиться и пройти от одного конца территории до другого, в начальном месте развития эпидемии рождается и подрастает значительное число восприимчивых детей. Таким путем эпидемия кори распространяется їв виде волн, обеспечивая непрерывность эпидемического процесса.
Еще недавно вирус кори был мало изучен и казался «одиноким» среди других вирусов, не имеющих родственных возбудителей. В настоящее время известны вирусы, близкие возбудителю кори по ряду биологических свойств (вирус чумы собак, а также другие вирусы этой группы).
Чума собак протекает у щенков как острая септическая инфекция, после выздоровления может наблюдаться состояние длительного носительства. У некоторых животных, особенно у взрослых собак, она может протекать в виде длительной латентной инфекции. Передается чума собак через выделения слизистых оболочек, и заражение происходит либо алиментарным путем, либо при контакте животных, в частности, потомство нередко заражается от матерей — латентных носителей вируса.
Изучение геномов вирусов кори и чумы собак методом секвенирования показало высокую степень их гомологии. В частности, это относится к гену нуклеопротеида, в котором определены области высокой (77%), средней (59%) и низкой степени гомологии [Rozenblatt S. et al., 1985]. Цитомегаловирусная болезнь чаще всего протекает как латентная инфекция, но нередко у маленьких детей в виде пневмоний или септических заболеваний с малохарактерными симптомами. Корь всегда имеет клинически выраженную картину и острое течение, сопровождающееся выработкой стойкого иммунитета, который сохраняется в течение всей жизни.
Мы провели параллель между тремя инфекциями не для того, чтобы утверждать, что цитомегаловирусная болезнь человека «произошла» от чумы собак, а корь является продуктом ее дальнейшей эволюции. Эта параллель проведена для того, чтобы показать весьма вероятный путь эволюции возбудителей многих инфекций человека. Если в случае оспы имеется адаптация возбудителя болезни домашних животных к организму человека с одновременным формированием острой инфекции, то в случае кори этот процесс проходил, по-видимому, в два этапа: сначала произошла адаптация паразита собак к организму человека, причем характер инфекционного процесса у человека остался таким же, как и у животного, или, может быть, инфекция стала еще более латентной; впоследствии произошло формирование острой инфекции типа оспы, каковой является современная корь. Можно лишь предполагать, почему в данном случае эволюция была двухступенчатой. Возможно, это произошло потому, что собака хотя и стайное животное, но контакт между стаями вряд ли был регулярным. Вирус чумы собак поэтому не мог сохраниться у этих животных, если бы не приобрел способности длительно латентно сохраняться в их организме. Естественно, что эти черты он сохранил, адаптировавшись к организму человека, и лишь в дальнейшем эволюционировал, став возбудителем кори — острой инфекции, при которой организм человека вырабатывает стойкий постинфекционный иммунитет, исключающий возможность носительства и длительного латентного сохранения вируса в организме.
Это могло произойти как путем постепенного накопления небольших изменений, так и путем мутаций. Корь могла появиться лишь в то время, когда общение между людьми стало интенсивным на больших территориях, насчитывающих многие миллионы человек, так как при иных условиях резервуар восприимчивых людей был бы недостаточным для сохранения возбудителя. Не надо забывать, что корью болеют один раз в жизни, а воздушно-капельный путь передачи и высокая восприимчивость приводят к тому, что каждая эпидемия этой болезни «подчищает» почти до нуля восприимчивую прослойку населения. Поэтому корь может существовать лишь непрерывно «блуждая» по большой территории. Когда же она снова «возвращается» на местность, где прежде была эпидемия, то к этому времени, рождается и подрастает достаточно детей, составляющих заметную прослойку восприимчивого населения. Но для этого нужны большие территории с достаточно плотным населением и выраженным постоянным общением между ними с тем, чтобы вирус кори мог «обойти» эту территорию за срок не менее 3—5 лет.
Эволюция кори шла таким образом, что она все более и более становилась эндемической детской инфекцией в густонаселенных районах земного шара, вызывая периодические, регулярно повторяющиеся эпидемии и оставаясь -заносной инфекцией для менее населенных и изолированных территорий, где поражаются все возрастные группы населения. В настоящее время корь — типичная инфекция развитого цивилизованного общества. Есть лишь одна возможность приостановить ее распространение — массовая иммунизация детей эффективной вакциной. Это мероприятие позволило почти сразу в 9—10 раз снизить заболеваемость корью. Дальнейшее изучение эффективности иммунизации против кори должно решить вопрос, будет ли эта инфекция ликвидирована, как это произошло с оспой, или же заболеваемость ею будет поддерживаться на достаточно низком уровне. По нашему мнению, задача ликвидации кори вполне реальная, но это требует интенсивных научных разработок.
По-видимому, сходный путь проделали и другие инфекции человека — вызванные парамиксовирусами 4 сероваров, а также респираторно-синцитиальным вирусом. Эволюция их далеко не закончилась и два из них (парамиксовирус 3 и респираторно-синцитиальный вирус) стали подлинным бедствием, особенно для детей.