Эволюция вирусов - Жданов В. М. 1990
Специальная часть
Бакуловирусы
Обширная группа бакуловирусов (семейство Baculoviridae) включает в себя вирусы ядерного полиэдроза и гранулеза насекомых. Будучи биологически обособлена от других вирусов, эта группа в то же время экологически не изолирована. Хотя большинство вирусов обнаружены у насекомых, сходные вирусы найдены у клещей, пауков, ракообразных и даже у грибов. Кроме того, несмотря на сходство представителей этой группы, условно объединяемой в семейство, имеются и значительные различия в подгруппах (родах), не говоря уже о вариациях величины геномов [Matthews R., 1982].
Геном представляет собой двунитевую циркулярную суперспирализованную ДНК с молекулярной массой 58х106—100х106, которая составляет 8—15% массы вирионов. Содержание Г+Ц колеблется в широких пределах — от 28% до 59%. Получены физические рестрикционные карты, указывающие, на сходство геномов некоторых изученных вирусов [Smith G. et al, 1979; Vlak J., Smith G., 1982].
В вирионах обнаружено до 15—25 белков, включая ферменты с молекулярной массой от 10 000 до 160 000. По крайней мере 8 белков выявлено в нуклеокапсидах, не менее 5 — во внешних оболочках [Tweeten К. et ah, 1980]. Мажорный белок нуклеокаисида VP12 сходен у вирусов полиэдроза и гранулеза и образует комплекс ДНК-белок, который сходен с гистонами [Tweeten К. et ah, 1980]. Весьма характерным является белок полиэдрин или соответствующий ему гранулин с молекулярной массой 25 000—33 000, он является главным компонентом телец включений. В образовании включений участвует еще 3 белка, а в вирионах имеется протеаза, разрушающая полиэдрин [Paune С., Ralmakoff J., 1978]. Между многими представителями семейства имеются антигенные связи [Summers М. et al., 1980; Smith G., Summers M., 1981].
Вирионы представляют собой один или несколько палочкообразных нуклеокапсидов, заключенных в единую оболочку. Размеры нуклеокапсидов колеблются в пределах 40—60х200—400 нм. Описаны бакуловирусы, в частицах которых количество ДНК варьирует в широких пределах [Krell Р., Stoltz D., 1979], имеются частицы с линейной ДНК [Odindo М, et al., 1986]. Возможно, в этих случаях речь идет о дефектных вирионах или о вирусах, не относящихся к рассматриваемой группе. Вирусы реплицируются в ядрах (подгруппы А и С), некоторые — частично в цитоплазме (подгруппа В). Окклюзия вирионов, столь характерная для вирусов этой группы, наступает в заключительной стадии, но может и не наступить (подгруппы С и D). Полидисперсная ДНК может также свидетельствовать о мультипартитности некоторых бакуловирусов.
Единственный пока род Baculovirus делят на 4 подгруппы, что должно быть признано условным, так как некоторые различия между подгруппами могут стать основанием для выделения их в отдельные роды или даже семейства. В подгруппу А входят вирусы ядерного полиэдроза, в одной оболочке могут содержаться несколько нуклеокапсидов (MNPV — multiple nucleocapsid particle virions) или один нуклеокалсид (SNPV — single etc). Вирусы полиэдроза Autographa californica и Bombyx mori являются представителями этих двух подтипов, поражающих насекомых и ракообразных (более 10 вирусов). К подгруппе В относятся вирусы гранулеза. Выделение их относительно условно, поскольку основные белки (полиэдрин и гранулин) имеют общие антигенные детерминанты. Более 50 вирусов этой подгруппы описаны у чешуекрылых. В подгруппу С включены вирусы, имеющие оболочечные нуклеокапсиды. В подгруппу D входят вирусы с полидисперсным геномом, причем тип D1 представлен вирионами с двумя внешними оболочками, а тип D2—множественными нуклеокапсидами, включенными в единую оболочку.
Вероятно, здесь нет надобности повторять общие соображения, высказанные при описании иридовирусов. Отметим лишь, что примат морфологии вряд ли может быть решающим для анализа этого достаточно вариабельного семейства, в котором тип D2 сближается с семейством полиднавирусов. Однако и в пределах этой группы предприняты попытки понять эволюцию отдельных представителей. С этой целью были сравнены некоторые параметры нескольких представителей рассматриваемой группы вирусов — аминокислотный состав полиэдрина и аминокислотная последовательность, частота встречаемости отдельных кодонов, степень гомологии, гидрофильность и гидрофобность полипептидных цепей.
На основании этих данных было составлено филогенетическое древо для 10 вирусов, поражающих чешуекрылых, двукрылых и перепончатокрылых (рис. 55). Приведенные данные указывают на возможность существования общего предка или немногих общих предков (по крайней мере у группы бакуловирусов). Если датировать их появление временем расцвета насекомых, то речь может идти о девоне (300—400 млн лет назад). На самом же деле время их появления следует датировать гораздо более поздними периодами, быть может, даже десятками миллионов лет назад, когда появились близкие к современным чешуекрылым перепончатокрылые и двукрылые. О сравнительно позднем появлении бакуловирусов свидетельствуют не слишком выраженная дивергенция генов для полиэдрина и гранулина, а также сохранение антигенных связей между некоторыми бакуловирусами. Не следует также забывать, что указанные группы насекомых являются быстро прогрессирующими и поэтому даже сопряженная эволюция вируса с «хозяином» могла явиться мощным стимулом видообразования, не говоря уж о завоевании новых экологических ниш.
При инфекции бакуловирусом личинок Autographa californіса и Trichopulsia nі выделен мелкий РНК-содержащий вирус, сходный с вирусом Nudaurelia capensis, а также с калицивирусами. Мажорный белок вирионов имеет молекулярную массу 67 000 [Morris Т. et аl., 1979].
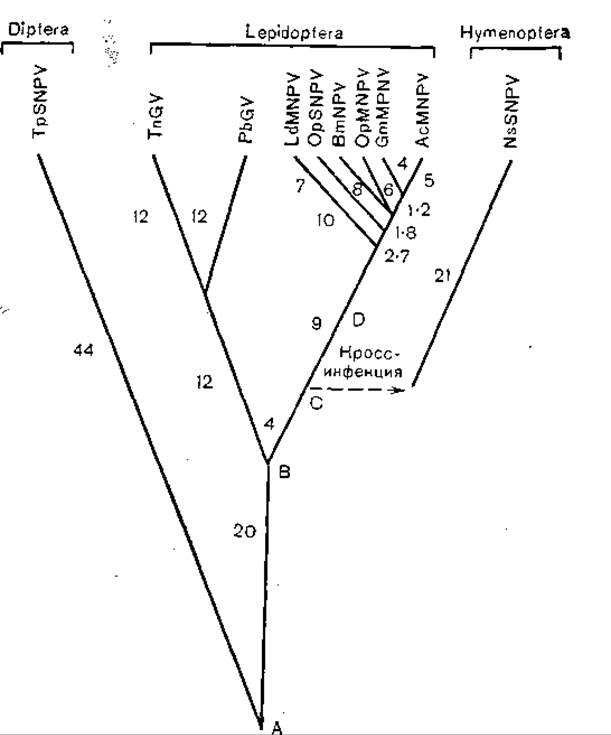
Рис. 55. Молекулярная филогения окклюзионных белков бакуловирусов. Цифры показывают процент различия в аминокислотном составе; пунктиром показано формирование бакуловирусов перепончатокрылых; А, В, С и D — точки дивергенции.