Эволюция вирусов - Жданов В. М. 1990
Специальная часть
Вироиды
Болезни, вызываемые вироидами, не имеют существенных отличий от других вирусных болезней растений. Первой из них, привлекшей внимание вирусологов, явилась веретеновидность, клубней картофеля (ВКК), позже были обнаружены болезни цитрусовых (экзокортис цитрусовых) и др. [Diener Т., 1982]. Возбудители этих болезней привлекли к себе внимание потому, что ими оказались не обычные вирусы, а сравнительно, небольшие, ковалентно замкнутые молекулы однонитевой РНК с молекулярной массой 100 000—120 000. У вироида ВКК она состоит из 359 рибонуклеотидов. Замыкание в кольцо РНК и. сложность вторичной структуры обеопечивают необычную устойчивость вироидов. На такой небольшой молекуле РНК не может быть закодирована даже «мелкая» белковая молекула, а лишь, может 'быть, короткий полипептид. На самом- деле РНК вироидов не имеет никакого генетического кода и даже кодонов АУГ.
Естественно, что вироиды не имеют собственных ферментов репликации РНК, она должна осуществляться клеточными ферментными системами. Как это происходит, пока неясно, хотя логично предположить существование репликативного интермедиата в виде матрицы РНК, комплементарной вирионной или двунитевой РНК. Такая двунитевая РНК была выделена из зараженных PSTV томатов с помощью электрофореза в полиакриламидном геле и ДНК — РНК-гибридизации. На циркулярной или линейной нити репликативного интермедиата обнаружены множественные вити вновь синтезируемой дочерней РНК Таким образом, моделью репликации РНК вироидов может служить вращающийся круг [Owens R., Diener Т., 1982]. Нельзя, однако, исключить и другой возможности — репликации через ДНК-интермедиат, о чем косвенно свидетельствует чувствительность ее к актиномицину D.
В этом отношении представляет определенный интерес изучение вироида болезни каданг-каданг, поражающей кокосовые пальмы на Филиппинах. При изучении этой болезни были обнаружены 4 вида низкомолекулярных РНК, состоящие из 250 и 500 нуклеотидов (ранние стадии болезни), 300 и 600 нуклеотидов (поздние стадии). По-видимому, первые члены пар являются мономерами, вторые — димерами первых. Обнаружена высокая степень гомологии этих вироидов с другими известными вироидами (PSTV, CSV, CEV, ASDV). К ним примыкают вирусоиды — инкапсидированные вирусоподобные РНК (VTMOV, SNMY, Subterranean clover mother virus). Все они не содержат генетического кода. Предполагают, что репликация их производится клеткой через репликативный интермедиат — олигомерную РНК [Haseloff J. et al., 1982].
Как уже указывалось, вторичная структура вироидов весьма сложна и включает в себя многочисленные комплементарные участки, что также делает возможным формирование димеров [Diener Т., 1986]. По результатам секвенирования у вироидов предположено существование 5 доменов: консервативный центральный район, район патогенности, домен вариабельный, два терминальных домена, взаимно обменивающихся между вироидами. Реаранжировка доменов иллюстрируется на примере вироида болезни каданг-каданг кокосовой пальмы, который появляется de novo при каждой инфекции. Этот механизм объясняет происхождение вироидов и вирусов, когда осуществляется обмен между РНК патогена и «хозяина» [Keese Р. et al., 1985]. На рис. 4 и 5 показано строение доменов вироидов, приведенных в табл. 4.
Таким образом, вироиды являются автономными фрагментами (их не хочется даже назвать генами) генетического материала клеток, транскрибированного в однонитевую РНК. Сложность их вторичной структуры обеспечивает защиту вироидов от нуклеаз и возможность молекулярной эволюции. До сих пор не ясно, что лежит в основе патогенеза вызываемых ими заболеваний. Скорее всего это не первичное свойство генетических элементов, от которых произошли вироиды, а приобретенное свойство, возможно, создавшее этим автономным структурам эколого-эволюционные преимущества. Имеются веские основания предполагать, что вироиды происходят от нитронов или транспозонов — подвижных генетических элементов. В пользу этой гипотезы свидетельствует структурное сходство вироидов и транспозонов. При сравнении 4 вироидов — карликовости верхушки томата (ТАSV), экзокортиса цитрусовых (CEV), веретеновидности клубней картофеля (PSTV), «планта махо» томата (TPMV). и задержки роста хризантем (CSV) — обнаружены 5 сходных последовательностей размерам 11—15 нуклеотидов, в целом их гомология составляет 73—83%.
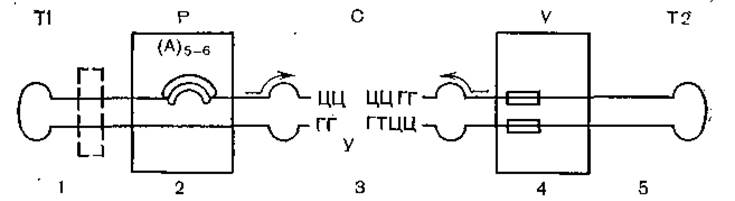
Рис. 4. Структура вироида (схема).
Показаны 5 участков гомологии между вироидами; стрелками показаны инвертированные повторы; 1 — левосторонний концевой участок; 2 — участок патогенности; 3 — консервативный центральный нуклеоид; 4 — вариабельный участок; 5 — правосторонний концевой участок.
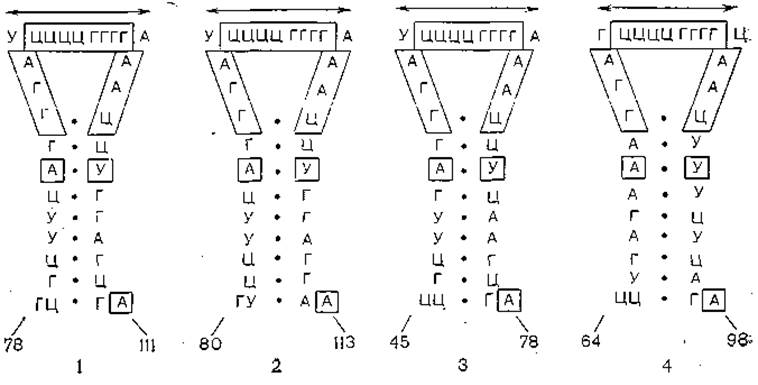
Рис. 5. Вторичная структура центрального консервативного участка различных вироидов.
Объединенные последовательности консервативны во всех вироидах; 1 —PSTV, TPMV: 2 — CEV, TASV; 3 — CCCV; 4 - HSV.
При этом обнаружены последовательности, типичные для концов транспозонов, а также инвертированные последовательности, оканчивающиеся динуклеотидами УГ и ЦА. На основании этих данных предполагается [Kiefer М. et al., 1983], что вироиды происходят от транспозонов или ретровирусных провирусов, утративших внутренние кодирующие- области. Выдвинута гипотеза, согласно которой вироиды, представляющие собой ковалентно-замкнутые кольцевые молекулы РНК с молекулярной массой 1,1 х 105—1,3 х 105, произошли в результате кольцевания отрезанных интронов. Такая модель выдвинута на основании изучения вироида веретеновидности клубней картофеля [Diener Т., 1986; Dinter-Gottlieb F., 1986].
Таблица 4. Вироиды и их изоляты с установленной нуклеотидной последовательностью
Хозяин |
Вироид |
Число нуклеотидов |
Авокадо |
АSBV |
247 |
Хризантема |
CSV |
354; 356 |
Цитрусовые |
CEV |
370—375 |
Орех |
CCCV |
246; 247 |
Огурец |
HSV |
297; 303 |
Картофель |
PSTV |
359 |
Томат |
TASV |
360 |
Томат |
TPMV |
360 |
Орех |
Болезни каданг-каданг |
246, 247 |
Авокадо |
кокосовой пальмы «Солнечной» пятнистости авокадо |
247 |
Хризантема Картофель |
Задержки роста хризантем Веретеновидности клубней картофеля |
359 |
Томат |
Карликовости верхушки томата |
360 |
«Планта махо» томата |
- |
|
Огурец |
Пожелтения плодов огурца |
297, 303 |
Цитрусовые |
Экзокортиса цитрусовых |
370—375 |
С другой стороны, вироиды по ряду свойств сближаются с некоторыми сателлитными вирусами, а обе группы структур — с нитронами ядерной и митохондриальной ДНК [Gollmer С. et al., 1985]. Цитированные авторы изучили сателлит кукумо-вируса — Peanut stunt virusassociated RN Ab (PARNAb). Его линейная РНК имеет на 5'-конце кэп-структуру и на 3'-конце труппу —ОН; 5'- и 3'-концы его гомологичны РНК другого сателлита вируса мозаики гороха. При этом до 90% нуклеотидов PARNA5 гомологично некоторым вироидам растений, а также некоторым нитронам (рис. 6). Эти данные позволяют заключить, что PARNA5 и вироиды происходят от нитронов ядерной и митохондриальной ДНК и эволюционно дивергировали.
Сателлиты вирусов растений имеют небольшую РНК, от 400 нуклеотидов и более, размножаются в присутствии хелпер- вирусов групп кукумовирус, неповирус, собемовирус, томбус-вирус. РНК-сателлит вируса кольцевой пятнистости табака имеет молекулярную массу 115 000 и состоит из 352 нуклеотидов. Инкапсидируется только плюс-нить в мономерной или мультимерной форме [Gerlach W. et al., 1986]. Вирус-хелпер относится к неповирусам и обеспечивает не только репликацию сателлитной РНК, но и ее инкапсидацию, причем вирус- сктеллит снижает патогенные свойства поддерживающего его вируса. Димерные формы РНК при автолизе образуют две активные молекулы PHК-сателлитов [Buzayan J. et al., 1986].
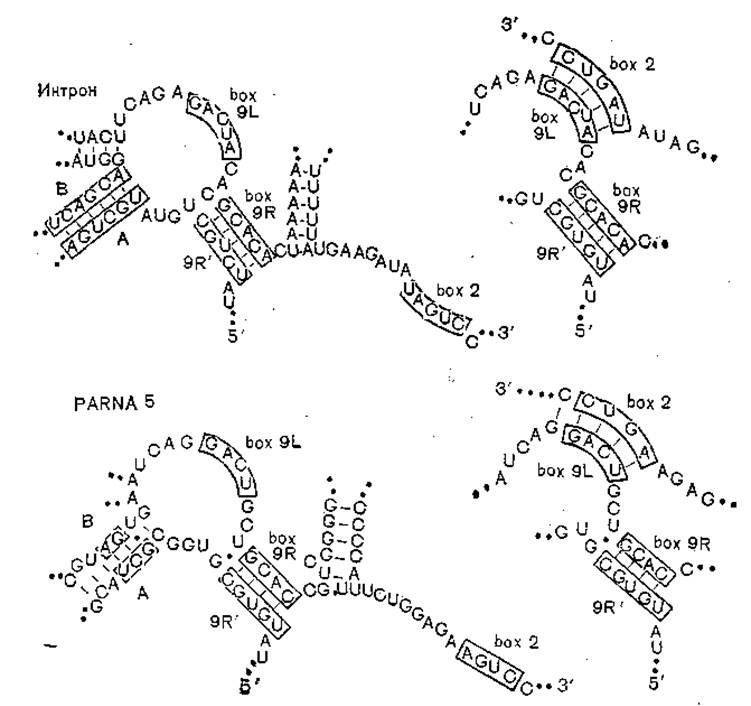
Рис. 6. Схематическое изображение возможного спаривания консервативных последовательностей интронов PARNA5 в сравнении с нитронами митохондрий. В рамку включены консервативные последовательности.
Наряду с вирусами-сателлитами РНК-содержащих вирусов часто «сопровождают» сателлитные РНК/ Так, провирус кольцевой пятнистости табака ассоциируется с кольцевидной сателлитной РНК, которая реплицируется основным вирусом по типу «катящегося круга» (rolling circle). Подобного рода сатeллитные РНК получили обозначение вирусоидов, часто обнаруживаемых в зараженных вирусами растениях [Lin- tliorst Н., Kapar J., 1984]. Сателлитная РНК (РНК2), выявляемая в растениях, пораженных cобемовирусами, отличается от их РНК (РНК1), которая имеет линейное строение и молекулярную массу 1,4 x 106. В отличие от них сателлитная РНК небольших размеров, обладает циркулярным строением и сходна с вироидами [Jones A., Mayo М., 1984].
Вирус-сателлит некроза табака размножается в присутствии основного вируса, однако кодирует собственный капсидный белок. В то время как вирус некроза табака имеет коэффициент седиментации 130S, вирус-сателлит осаждается при 58S. Серологические белки обоих вирусов не имеют никакого родства; размер вирионов около 30 нм, сателлита— 17 нм. Сателлитная РНК имеет молекулярную массу 0,4 х 106, что соответствует 1200 нуклеотидам [Kassanis В., 1981]. При изучении сателлитной РНК вируса мозаики огурца была показана большая степень ее гомологии с 3'-концевой частью тРНК и соответствующих генов растения [Gordon К., Symons R., 1983].
Выявлены своеобразные соотношения между квадрапартитным вирусом хронического паралича пчел и его сателлитом. Их капсидные белки разные и серологически не имеют родства. Вирус хронического паралича пчел является хелпером ассоциированного с ним вируса-сателлита, не способного к самостоятельной репродукции. При совместном заражении происходит конкуренция за использование полимеразы, в связи с чем титр вируса-хелпера снижается [Ball В. et al., 1985].
Таким образом, вироиды и вирусоиды могут рассматриваться как самые начальные стадии образования автономных генетических структур, из которых впоследствии могут развиться классические вирусы. Их происхождение от клеточных генетических элементов несомненно, вместе с тем уже на этой первоначальной стадии они характеризуются автономией эволюции.