ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 1 - Г.Й. Щербак - 1995
ПІДЦАРСТВО БАГАТОКЛІТИННІ (METAZOA)
РОЗДІЛ СПРАВЖНІ БАГАТОКЛІТИННІ (EUMETAZOA)
ТИП РЕБРОПЛАВИ (CTENOPHORA)
КЛАС ТРЕМАТОДИ, АБО ДИГЕНЕТИЧНІ ПРИСИСНІ (TREMATODA, АБО DIGENEA)
Усі трематоди — ендопаразити. Дорослі особини (марити) трапляються переважно в різних відділах травного тракту хребетних, а також у легенях, нирках, порожнині тіла, кровоносній системі хребетних тварин. Серед них є багато збудників тяжких хвороб людини та сільськогосподарських тварин. Розміри трематод коливаються від 0,3— 0,4 мм до 3—7,6 см. Відомо понад 4 тис. видів, в Україні зареєстровано близько 600 видів.
Форма тіла тварин найчастіше листоподібна. У дорослих червів, що називаються маритами, є, як правило, два добре розвинені присоски. Один із них (ротовий) розташований на передньому кінці тіла й у центрі містить ротовий отвір, другий (черевний) функціонує лише як орган прикріплення. Звичайно він розташований в центрі тіла, проте у деяких видів він зміщений ближче до переднього або заднього кінця. Ступінь розвитку присосків залежить від місця локалізації паразита.
У трематод, які паразитують у кишковому тракті, присоски міцні, а у тих, що мешкають у порожнині тіла або кров'яному руслі — недорозвинені чи їх зовсім немає.
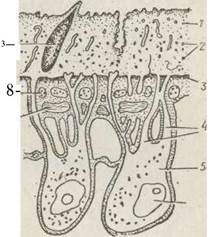
Рис. 143. Схема будови покривів трематод (за даними електронної мікроскопії):
1 — зовнішня без'ядерна цитоплазматична пластинка; 2 — мітохондрії; 3 — базальна мембрана; 4—цитоплазматичні тяжі, що з'єднують зовнішню і занурену частини епітелію; 5 —занурені ділянки цитоплазми з ядрами (5); 7 — кільцеві м'язи;
8— поздовжні м'язи; 9 — кутикулярний шипик
Покрив трематод, як і всіх інших паразитичних червів, на відміну від вільноживучих турбелярій, позбавлений війок і є тегументом, побудованим за типом зануреного епітелію, (рис. 143). Зовнішні частини епітеліальних клітин зливаються своїми бічними поверхнями й утворюється синцитій, тобто цитоплазматична пластинка без клітинних перегородок. Від неї відходять цитоплазматичні відростки з ядрами, які досягають паренхіми. Цитоплазматична пластинка підстелена базальною мембраною, яка переривається в місцях відгалуження цитоплазматичних відростків. Під мембраною залягають кільцевий та поздовжній шари м'язів.
Травна система трематод, як правило, добре розвинена й подібна до травної системи турбелярій, проте в деяких дрібних форм спостерігається її рудиментація. Травна система починається ротовим отвором, що веде в мускулясту глотку, яка переходить у стравохід. Стравохід продовжується у дві сліпо замкнені гілки середньої кишки.
У великих за розміром трематод, наприклад Fasciola hepatica, гілки середньої кишки розгалужені, що пов'язано з їх транспортною функцією, всі відгалуження закінчуються сліпо (рис. 144).
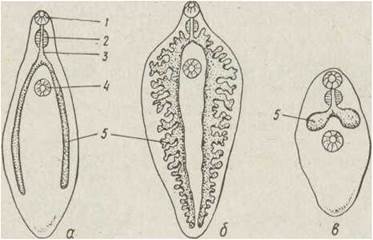
Рис. 144. Травна система трематод — Opisthorchis (a), Fasciola (б), Gymnophalus (в): 1 — ротовий отвір; 2 — глотка; 3 — стравохід; 4 — черевний присосок; 5 —середня кишка
Видільна система тварин належить до протонефридіального типу, головні збираючі канали впадають у сечовий міхур, що відкривається в зовнішнє середовище екскреторною порою.
Нервова система трематод — типовий ортогон. Вона Укладається з ортогонного мозку, від якого вперед відходять нервові стовбури, що іннервують ротовий присосок і, розгалужуючись, заходять у зовнішній цитоплазматичний шар тегументу переднього кінця тіла. Назад від мозку напрямлені два вентральні (найтовщі) стовбури, два дорзальні та два латеральні. Всі поздовжні стовбури з'єднані між собою поперечними комісурами, що мають вигляд кілець або напівкілець. Більшість їх розташована в передній частині тіла. Органи чуття в дорослих форм розвинені слабо, що пов'язано з паразитичним способом життя.
Статева система марити гермафродитна. Виняток становлять роздільностатеві кров'яні сисуни родини Schistosomatidae.
Чоловіча статева система представлена двома, рідше кількома сім'яниками різної форми — від округлих до деревовидно розгалужених.
Від сім'яників ідуть сім'япроводи, що зливаються в спільний канал, який починається потовщенням — сім'яним міхурцем. Цей міхурець переходить у сім'явипорскувальний канал, що пронизує копулятивний орган. Він відкривається в статеву клоаку й може випинатися назовні (рис. 145).
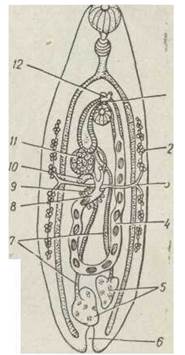
Рис. 145. Схема будови статевої системи марити трематод:
1 — статева клоака; 2 — жовтівники; 3 — жовтівникові протоки; 4— матка; 5 — сім'яники; 6 — сечовий міхур; 7 — сім'япроводи; 8— оотип; 9 — тільце Меліса; 10 — сім'яприймач; 11 — яєчник; 12 — копулятивний орган
Жіноча статева система складається з одного яєчника різної форми. Короткий яйцепровід веде до невеликого мішковидного утвору — оотипу. В нього впадають протока сім'яприймача, численні одноклітинні залози, що утворюють тільце Меліса, та канал резервуара, в якому накопичується продукція пари жовтівників. Від оотипу відходить лаурерів канал, що відкривається на спині, та матка у вигляді звивистої довгої трубки, яка виходить у статеву клоаку. Під час копуляції копулятивний орган однієї особини вводиться в матку іншої, по ній сперматозоїди через оотип потрапляють до сім'яприймача, де зберігаються протягом усього життя. Власне яйцеклітини запліднюються в оотипі, куди вони одна за одною надходять із яєчника; сюди ж потрапляють із сім'яприймача невеличкими порціями сперматозоїди, а також жовточні клітини. Запліднена яйцеклітина оточується жовточними клітинами та вкривається міцною оболонкою, яка утворюється з секрета тільця Меліса та жовтівників; сформовані яйця виходять у матку та просуваються по ній до вивідного отвору, поступово дозріваючи. Невикористані на запліднення сперматозоїди та надлишки жовточних клітин виводяться через лаурерів канал назовні. Трематоди дуже плодючі, в середньому кожні 20—30 с продукується нове яйце.
Усі трематоди мають дуже складний життєвий цикл, що супроводжується чергуванням поколінь, паразитичних і вільноживучих фаз розвитку та зміною хазяїв (рис. 146).
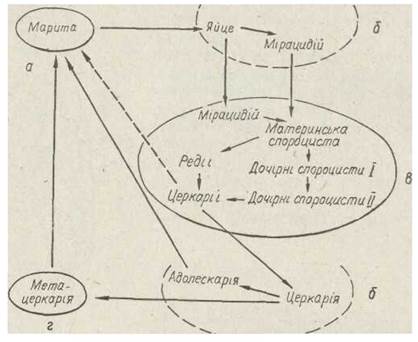
Рис. 146. Узагальнена схема життєвого циклу трематод — паразит у остаточному хазяїні (а), у зовнішньому середовищі (б), у першому проміжному хазяїні (в), у другому проміжному хазяїні (г)
Як уже згадувалося, марити паразитують у внутрішніх органах хребетних тварин. Яйця, що їх продукують паразити, в типовому випадку мають потрапити у воду, де з них виходять рухливі личинки — мірацидії (рис. 147). Тіло» мірацидія, як правило, вкрите великими плескатими війчастими клітинами (епітеліальними пластинками), в цитоплазмі яких є великі запаси глікогену, що служить енергетичним матеріалом, необхідним для роботи війок. Під епітеліальними пластинками містяться гіподерма, шари кільцевих і поздовжніх м'язів. Травної системи у мірацидіїв; немає, вони не живляться, існують за рахунок запасів поживних речовин. Видільна система представлена двома протонефридіями з двома—чотирма циртоцитами, що відкриваються самостійними порами. Мірацидії мають мозковий ганглій, який іннервує добре розвинені органи чуття — є пара інвертованих очей та сенсили, очевидно, різного призначення (хеморецептори, тактильні тощо).
На передньому кінці тіла міститься хоботок, що відіграє важливу роль у процесі проникнення мірацидія в тіло першого проміжного хазяїна, яким для трематод є молюск певного виду, переважно з класу Черевоногих. На верхівці хоботка відкриваються протоки апікальної залози, секрет якої руйнує покриви молюска, що забезпечує проникнення мірацидія. Апікальну залозу називають органом проникнення. У задній частині тіла мірацидія містяться зародкові клітини. Протягом короткого часу мірацидії мають знайти першого проміжного хазяїна і проникнути в нього. У цей період велику роль відіграють таксиси, що полегшують їх зустріч. Так, для мірацидіїв багатьох видів характерні негативний геотаксис і позитивний фототаксис, їх поєднання приводить до того, що мірацидії збираються біля поверхні води, куди регулярно піднімаються для, дихання легеневі молюски — проміжні хазяї цих трематод. Важливу роль відіграють реакції личинок на хімічні подразнення. У багатьох видів трематод (Opisthorchis felineus, Dicrocoelium dendriticum) мірацидії не виходять із яйця у зовнішнє середовище, й зараження молюсків відбувається під час поїдання ними яєць, що містять цілком сформовані личинки.
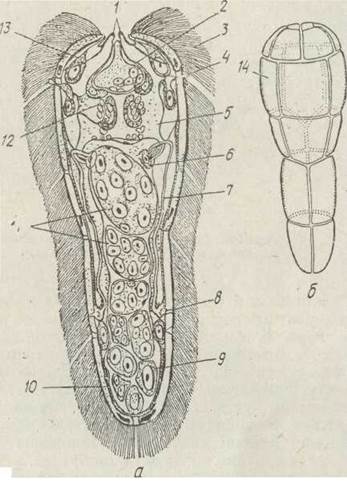
Рис. 147. Будова мірацидія — мірапидій Fasciola hepatica (а), схема розміщення епітеліальних пластинок (б): 1 — втягнутий хоботок; 2 — апікальна залоза; 3 —війки; 4— сенсили; 5 — мозковий ганглій; б — циртоцит; 7 — канал протонефридія; 8 — сечовий міхурець; 9 — генеративні (зародкові) клітини; 10 — м’язовий шар; 11 - зародки, що розвиваються; 12 — пігментне очко; 13 — залозиста клітина; 14 — епітеліальні пластинки
Після проникнення в тіло молюска мірацидій перетворюється на статевозрілу материнську спороцисту, яка. має значно простішу будову, ніж мірацидій. У неї зникають очі, хоботок, апікальна залоза тощо: Личинка втрачає епітеліальні пластинки, й гіподерма, що знаходиться під ними, стає тегументом спороцисти, тобто має місце регресивний метаморфоз. Тіло в спороцисти мішкоподібне, округле або видовжене, червоподібне. У деяких видів трематод спороцисти розгалужені (рис. 148). Спороциста не має кишкового тракту і живиться всією поверхнею тіла. Нервова система та органи чуття в неї майже не розвинені.
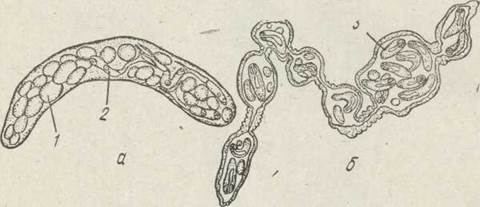
Рис. 148. Мішкоподібна (а) та нитчаста (б) спороцисти:
1 — зародкові кулі; 2 — церкарії; 3 — редії
Материнська спороциста розмножується партеногенетично. Партеногенетичні яйця, якими заповнене тіло спороцисти, без запліднення починають дробитися, утворюючи ембріони, та-к звані зародкові кулі, з яких формується друге партеногенетичне покоління трематод, представлене двома різними формами — редіями або дочірніми спороцистами. Типові редії мають витягнуте циліндричне тіло з парою локомоторних виростів, що розташовані в задній третині тіла. У редій добре розвинений шкірно-м'язовий мішок, є травна система, що починається ротовим отвором, який веде в глотку і далі в довгий мішкоподібний кишечник. Протонефридії в них парні, кожен із них має власну екскреторну пору; нервова система типу ортогону, але органи чуття розвинені погано (рис. 149).
Дочірні спороцисти, на думку вчених,— це неотенічні редії, тобто редії, що починають розмножуватися на ранніх етапах розвитку.організму. Дочірні спороцисти мають мішкоподібну, червоподібну або округлу форми, травної системи в них немає. На відміну від материнської спороцисти дочірня має, як і редії, «пологову» пору, через яку виходить наступне покоління трематод — церкарії.
Вихід редій і дочірніх спороцист, які досить рухливі в молодому віці, з материнської спороцисти відбувається у разі їх одночасного дозрівання через розрив її стінок або в процесі поступового формування особин через невеликі розриви стінок, які швидко затягуються й спороциста продовжує жити та розмножуватися.
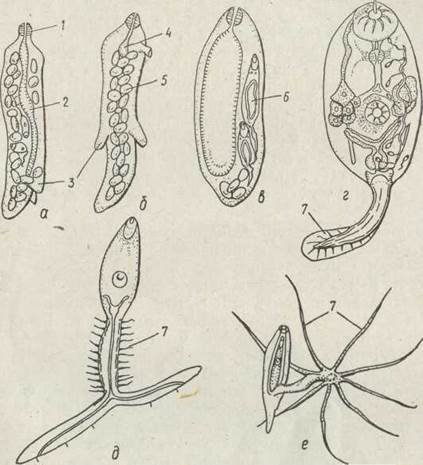
Рис. 149. Будова редій та церкарій — типові редії (а, б), мішкоподібна редія (в), церкарії (г, е): 1 — глотка; 2 — кишечник; 3 — локомоторні вирости; 4 — рудиментарний кишечник; 5 —зародки церкарій; 6 —сформовані церкарії; 7 — хвостовий придаток
Редії та дочірні спороцисти локалізуються в порожнині тіла молюска, прикріплюються до поверхні його кишечника та інших внутрішніх органів, але найчастіше вони паразитують у печійці. Редії активно живляться тканинами печінки й, крім того, можуть поглинати поживні речовини всією поверхнею тіла. Спороцисти харчуються лише через покриви. Тегумент спороцисти вкритий численними, мікроворсинками, спеціальні залозисті клітини секретують всі необхідні травні ферменти, які перетравлюють ділянки тканини, що щільно прилягають до стінки спороцисти, тобто має місце пристінкове травлення з тією різницею, що відбувається воно не в організмі, а поза ним.
Розмноження редій та дочірніх спороцист відбувається партеногенетично з зародкових клітин, які, як і в материнської спороцисти, розвиваються в порожнині їх тіла. Із зародкових клітин можуть формуватися дочірні редії та спороцисти або одразу церкарії. Продуктивність редій не дуже велика (дві-три дочірні редії або шість — десять сформованих церкарій), а нащадків спороцисти налічується сотні тисяч. Це зумовлено поліембріонією, тобто розвитком із однієї яйцеклітини кількох, а іноді й багатьох зародків.
Повністю сформовані церкарії за своєю будовою близькі до марит, від яких відрізняються меншими розмірами та наявністю мускулястого хвоста різної будови (див. рис. 149). У них є присоски, майже сформована травна система (яка, проте, не функціонує), протонефридіальна видільна система та добре розвинена нервова система. Органи чуття представлені численними сенсилами, що сприймають хімічні та механічні подразнення. Церкарії мають різноманітні залози, ступінь розвитку яких залежить від їх біології. Так, у личинок, що інцистуються у зовнішньому середовищі, найбільше розвинені цистогенні залози, їх секрет витрачається на побудову товстої оболонки цисти. У церкарій, що паразитують далі в другому проміжному хазяїні, ці залози розвинені гірше. Дуже розвинений комплекс залоз проникнення, секрет яких допомагає проникнути через покриви хазяїна. Є ще слизові залози, їх секрет вкриває тіло церкарії чохликом, захищаючи від дії ферментів молюска, коли церкарія мігрує в його тілі.
Церкарії деяких видів не потребують другого проміжного хазяїна. Одні з них осідають на певний субстрат і там інцистуються, перетворюючись на адолескарію. Вона вкрита товстою оболонкою з двох-чотирьох шарів, під її захистом майже нерухома адолескарія дуже повільно втрачає запаси глікогену й тому зберігає життєздатність протягом тривалого часу (але не росте й не розвивається), поки її не з'їсть остаточний хазяїн. Інші церкарії активно проникають у тіло остаточного хазяїна.
У багатьох видів подальший розвиток відбувається в другому проміжному хазяїні (риби, личинки водних комах, інші молюски, ракоподібні тощо). Церкарії протягом 24— 48 год (час, за який витрачається глікоген, що запасається в тілі личинки, зокрема в хвостовому придатку) мають зустрітися з другим проміжним хазяїном. Як і у мірацидіїв, у церкарій є ряд адаптацій (гео-, фото- та хемотаксиси), що полегшують пошук хазяїна та проникнення в нього.
Проникнення через покриви нового хазяїна, у різних видів відбувається по-різному. В одних церкарій, наприклад, є сильний кутикулярний стилет, яким покриви розрізаються, у інших — короткий міцний хоботок із гачками, якими травмуються пркриви, а в ранки вводиться секрет залози проникнення. Місце проникнення визначається за допомогою сенсил, що сконцентровані на передньому кінці тіла церкарії. Проникаючи в тіло хазяїна, церкарії відкидають хвіст. У другому проміжному хазяїні вони локалізуються в м'язах або внутрішніх органах, де церкарії інцистуються, перетворюючись на метацеркарій. Поряд з активним існує також пасивне проникнення церкарій у другого проміжного хазяїна. Так, личинки деяких трематод заковтуються рибою або засмоктуються разом із водою та інцистуються на стінках ротової порожнини. Є види, церкарії яких осідають на поверхні тіла равликів, а потім заповзають у легеневу порожнину, де й інцистуються. Цисти мають тонку захисну стінку, через яку можуть проникати поживні речовини. Личинка (метацеркарія), що міститься в цисті, росте, розвивається, проте метацеркарія (як і адолескарія) перетворюється на мариту лише в організмі остаточного хазяїна.
Таким чином, у життєвому циклі трематод чергуються одне гермафродитне (марита) та кілька партеногенетичних поколінь (спороцисти, редії). Такий цикл називається гетерогонією. Доросла особина гермафродитного покоління — марита, партеногенетичних ·—спороциста, редія. Мі- рацидій — це личинка спороцисти, а церкарія, метацеркарія, адолескарія — личинки марити.
Як уже згадувалося, серед трематод є велика кількість видів, що викликають небезпечні захворювання (трематодози) людини, домашніх і промислових тварин. Патогенний вплив трематод на організм хазяїна має різний характер. Паразити порушують цілісність тканин, викликають закупорку проток різних органів, негативно впливають на хазяїна через виділення токсичних продуктів обміну речовин тощо. У разі інтенсивних інвазій (уражень) спостерігається виснаження організму хазяїна. Трематоди можуть спричинювати розвиток різних новоутворень, зокрема й злоякісних.
Система класу Трематода нині розробляється. Вчені виділяють у класі від чотирьох до десяти рядів. Розглянемо життєві цикли найвідоміших збудників трематодозів.
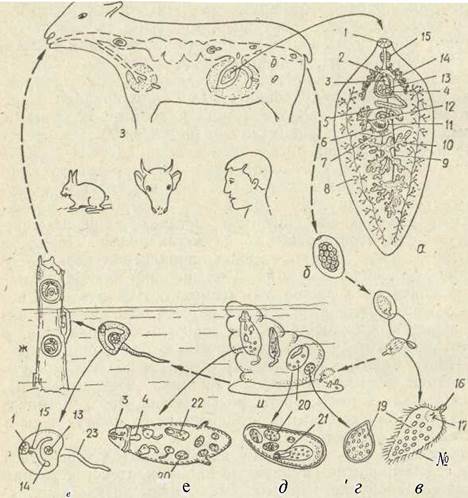
Рис. 150. Життєвий цикл Fasciola hepatica — марита (а), запліднене яйце (б) мірацидій (в), молода материнська спороциста (г) зріла спороциста (д), редія (е), церкарія (є), адолескарія на траві (ж ) основні хазяї (з), проміжний хазяїн молюск Limnea truncatura (и). 1 -ротовий присосок; 2 - статева клоака; 3 — копулятивний канал; 4 - сім'явипорскувальний канал; 5 — мичка; 7— токи; 8 — жовтівник; 9 — сім’япровід; 10 — сім’ яник; 11 — яйцепровід. 12 — яєчник; 13 — черевний присосок; 14 —кишечник; 15 —глотка; 17 — залоза проникнення; 19 — зародкові клітини; 20 — зародкові кулі, 21 - материнська редія; 22 - дочірня редія; 23 — хвіст
Печінковий сисун (Fasciola hepatica) — досить великий (26—3 см завдовжки) листоподібний паразит, що живе в жовчних протоках печінки овець, великої рогатої худоби, рідше інших тварин і людини. Ротовий та черевний присоски зближені, двогіллястий кишечник сильно розгалужений, як і статеві залози (рис. 150).
Марити F. hepatica дуже плодючі, протягом тижня одна особина продукує близько мільйона яєць, що виводяться через жовчні протоки та кишечник назовні. Розвиток яєць та вихід мірацидіїв відбувається тільки у воді. Перший проміжний хазяїн —малий ставковик Limnea truncatula, Після проникнення в тіло молюска мірацидій перетворюється на материнську спороцисту, яка утворює нове покоління — редії, що розривають материнську спороцисту, виходять у порожнину тіла молюска й потрапляють до травної залози. Тут народжується кілька поколінь, редій, а потім церкарії, які виходять у воду, недовго плавають, потім прикріплюються за допомогою присосок до водних рослин або до поверхневої плівки води, відкидають хвіст й інцистуються, перетворюючись на адолескарій. Інцистована аделескарія витримує висихання й зберігає життєздатність навіть у сухому сіні. Отже, людина може заразитися, п'ючи воду із стоячих водойм, а тварини — ще й поїдаючи рослини на заболочених пасовищах або сіно.
Фасциольоз — це поширена на земній кулі хвороба, проте у людей вона трапляється досить рідко.
Котячий сисун Opistnorchis felineus є збудником дуже небезпечної хвороби — опісторхозу (рис. 151). Паразит локалізується в жовчних протоках печінки, жовчному міхурі, іноді в протоках підшлункової залози людини та дуже широкого кола ссавців. Перший проміжний хазяїн — прісноводний молюск Bithynia leachi, другий — різні види коропових риб. Церкарії проникають під покриви риб та інцистуються в їх м'язах, перетворюючись на метацеркарії. Зараження відбувається внаслідок поїдання риби, яка недостатньо просолена, проварена чи просмажена. Проте найчастіше опісторхоз трапляється в районах, де використовують в їжу заморожену рибу, так звану струганину. Інтенсивність інвазії буває надзвичайно високою. Під час розтину людини, що загинула від опісторхозу, в її печінці було виявлено 25320 трематод. Опісторхоз є поширеною хворобою, зокрема в Україні.
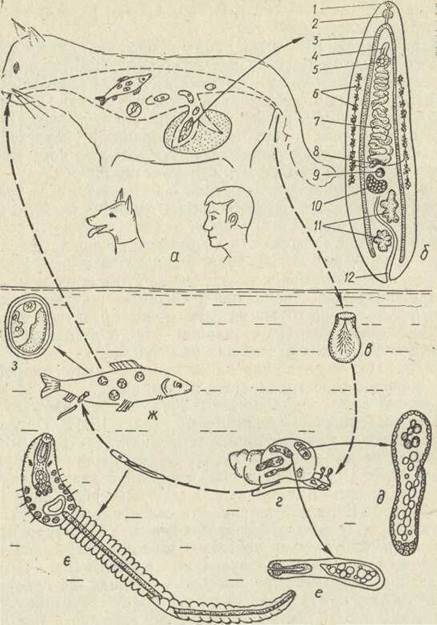
Рис. 151. Життєвий цикл Opisthorchis felineus — основні хазяї (а), марита (б), яйце з мірацидієм (в), перший проміжний хазяїн молюск Bittiynia leachi (г), спороциста (3), редія (е), церкарія (е), другий проміжний хазяїн (ж), метацеркарія (з): 1—ротовий присосок; 3 — глотка; 3 —кишечник; 4 — копулятивний орган; 5 — черевний присосок; б — жовтівники; 7—матка; 8 — жовтівникові протоки; 9 — сім'ядриймач; 10 — яєчник; 11 —сім'яники; 12 — екскреторний міхур
Ланцетоподібний сисун Dicrocoelium dentriticum є збудником дикроцеліозу. Він паразитує в жовчних протоках печінки овець, великої рогатої худоби, зрідка людини та багатьох диких тварин .(рис. 152). Розвиток цього сисуна не пов'язаний з водним середовищем. Яйця дикроцелія з мірацидіями потрапляють разом із екскрементами тварин на грунт. Вони вкриті дуже товстими оболонками, тому не висихають навіть протягом кількох місяців. Першим проміжним хазяїном паразита є наземні молюски (Helicella, Zebrina та ін,), які поїдають його яйця. У кишечнику молюска з яйця виходить мірацидій. Він проникає в печінку, де перетворюється на материнську спороцисту, всередині якої
розвиваються дочірні спороцисти, а з них одразу виникають церкарії (стадія редії не утворюється). Церкарії виходять у легеневу порожнину молюска, там вони вкриваються слизом, склеюються разом і виводяться через дихальний отвір молюска у вигляді «збірних цист», що містять тисячі церкаріїв.
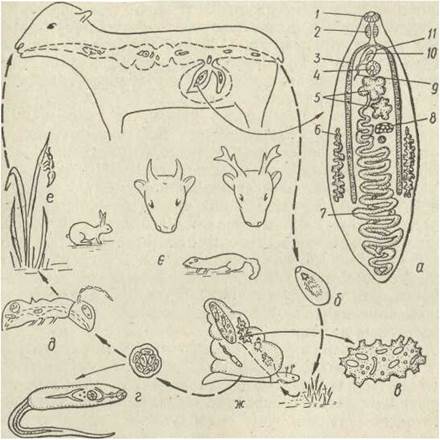
Рис. 152. Життєвий цикл Dicrocoelium dendriticum— марита (;і), яйце з мірацидієм (б), спороциста (е), церкарії у слизовій збірній цисті (г), метацеркарії в мозку та тулубі мурашки (д), другий проміжний хазяїн — мурашка (е), основні хазяї (е), перший проміжний хазяїн — наземний молюск (ж): 1 — ротовий присосок; 2— глотка; 3 — кишечник; 4 — черевний присосок; 5 — сім’яники; б — жовтівники; 7 — матка; 8—яєчник; 9—сім'явипорскувальний канал; 10 — копулятивний орган; 11 — яйцепровід
Другим проміжним хазяїном ланцетоподібного сисуна є мурашки роду Formica. Вони заносять «збірні цисти» до мурашників і поїдають, заражуючись дикроцелієм. Масовому зараженню мурашок сприяє те, що вони діляться своєю їжею з іншими особинами мурашника. У мурашки, яка з'їла «збірну цисту», одна з церкарій проникає у мозок,
перетворюючись на «мозкового черва», а всі інші — у мускулатуру, де перетворюються на метацеркарії. Під дією «мозкового черва» у заражених мурашок настає параліч, вони виповзають на траву, чіпляються щелепами за край листової пластинки й нерухомо висять на ній. Травоїдні тварини ковтають їх разом із травою. У кишечнику цих тварин мурашки перетравлюються, а метацеркарії звільняються з цист, проникають у жовчні протоки печінки й жовчний міхур і там перетворюються на марит. Людина може заразитися, випадково проковтнувши заражену мурашку, але це трапляється дуже рідко.
Специфічними паразитами людини є трематоди роду Schistosoma — Sch. haematobium і Sch. mansoni, що паразитують у кров'яному руслі хазяїна. Характерною особливістю цих трематод є роздільностатевість. Ширший за самку самець охоплює її загорненими на черевний бік краями тіла (рис. 153). Життєвий цикл шистосом пов'язаний лише з двома хазяями. Марити локалізуються в мезентеріальних венах (збудники кишкового шистосоматозу Sch. mansoni) або у венозних сплетіннях сечостатевої системи людини (Sch. haematobium). Яйця шистосом осідають,у капілярах ї проникають у товщу тканин. Тут завдяки м'язовим скороченням вони надходять до кишечника або сечового міхура й виводяться назовні.
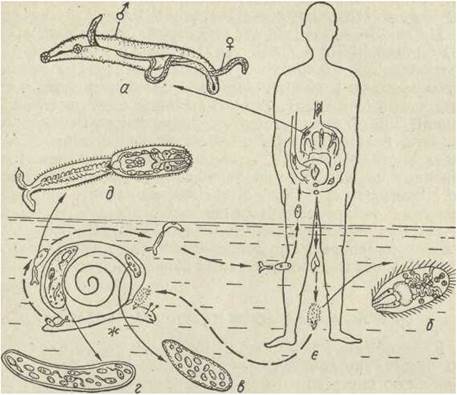
Рис. 153. Життєвий цикл Schistosoma haematobium:
а — статевозрілі самець і самка; б —мірацидій; е—спороциста; г — редія; д — церкарія; е — церкарія в позі очікування; є — основний хазяїн; ж — проміжний хазяїн
Для подальшого розвитку яйця мають потрапити у воду, де мірацидій відшукує молюска певного виду, в якому і відбуваються всі наступні перетворення на спороцисту, формування редій та церкарій. Церкарії мають позитивний фототаксис, завдяки чому підвішуються до поверхневої плівки води й у нерухомому стані («поза очікавання») перебувають кілька діб. У разі появи людини церкарії швидко переходять на неї. Зараження відбувається внаслідок активного проникнення крізь шкіру людини під час купання, роботи на рисових плантаціях тощо. У тілі людини шистосоми можуть жити кілька років. Вони спричинюють досить тяжкі захворювання сечостатевих органів.
Шистосоматоз людини — хвороба південних і тропічних країн. В Україні він не трапляється, проте в людей, які купаються в стоячих зарослих ставках або затоках річок, спостерігається шистосомний дерматит — захворювання шкіри, яке називають водяною сверблячкою. Ця хвороба спричинюється церкаріями тих видів шистосоми, які паразитують у водоплавних птахів: качок, мартинів тощо. Подразнення шкіри людини пов'язане з механічною дією личинок, що вбуравлюються в шкіру, та з дією речовин, які вони виділяють. У шкірі людини церкарії гинуть. Дерматит трапляється досить часто в Україні, Росії (дельта Волги), в деяких районах Казахстану, у США.
На закінчення слід згадати життєвий цикл присиснів із роду Leucochloridium, що паразитують у птахів. Яйця зі сформованими мірацидіями розсіюються птахами разом із екскрементами й потім поїдаються наземними молюсками- янтарками (Succinea). В їх кишечнику вилуплюються мірацидії, що мігрують у печінку, де перетворюються на спороцисти, в яких формуються безхвості церкарії. Спороциста росте, галузиться, її найдовші гілки, заповнені розвиненими церкаріями, проникають у щупальця молюска, які розтягуються. До того ж ці гілки спороцисти набувають яскравого забарвлення (зелене, буре, нерідко з білими смугами та червоно-коричневими плямами). Колір та пульсуючі рухи спороцисти добре видно крізь тонкі покриви щупалець, завдяки чому птахи їх охоче з'їдають (рис. 154).
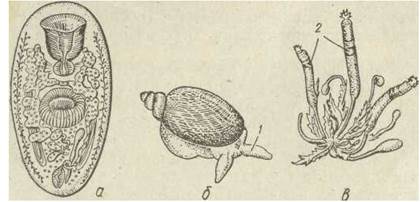
Рис. 154. Leucochloridium paradoxum— марита (а), заражений слимак янтарка Succinea (б), спороциста з печінки слимака (а):1—здуті щупальця молюска з виростами спороцисти; 2— пігментовані мішковидні вирости спороцисти