ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 2 - Г.Й. Щербак - 1996
ТИП КІЛЬЧАСТІ ЧЕРВИ (ANNELIDA)
КЛАС БАГАТОЩЕТИНКОВІ (POLYCHAETA)
Цей клас об'єднує понад 6 тис. видів морських (переважно) та прісноводних (рідше) тварин. Відомий один тропічний вид Lycastopsis catarractarum із родини Nereidae, який живе на плантаціях бананів і какао й живиться гнилим листям, солодкими плодами. У Чорному та Азовському морях знайдено близько 200 видів.
Розміри поліхет становлять від кількох міліметрів до кількох метрів (Eunice gigantea).
Тіло багатощетинкових (рис. 2) складається з головної лопаті (простомій), сегментованого тулуба та анальної лопаті (пігідій). На простомії розташовані органи чуття: пара пальп, пара або кілька пар антен, очі, нюхальні ямки. У різних представників певні органи простомія можуть видозмінюватись або зникати, особливо у риючих та сидячих форм. Простомій разом із першим або ще двома-трьома передніми сегментами (перистомієм) утворюють головний відділ. Злиття передніх сегментів, що веде до утворення головного відділу, зветься цефалізацією. На перистомії розташовані рот і вусики, які виконують функцію органів чуття.
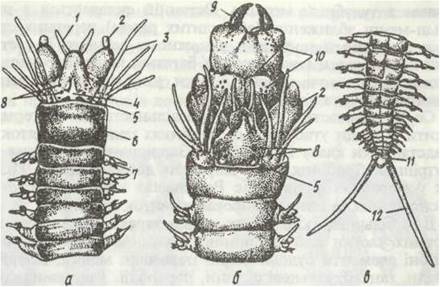
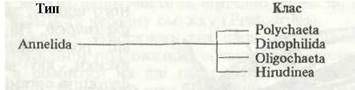
Рис. 2. Зовнішня будова Nereis pelagica:
а — передній кінець із втягнутим букальним відділом; б — з вивернутим; в — задній кінець; 1 - антена; 2 - пальп; 3 - вусики перистомія; 4 - простомій; 5 - нюхальна ямка; 6 - перистомій; 7-траподій; 8 -очі; 9- щелепа; 10 -букальний відділ; 11-тідій; 12 - пігідіальні вусики
Тулуб складається з різної кількості сегментів (від 5 до 800).
У більшості поліхет кожний сегмент тулуба, крім перистомальних, має по парі бічних виростів — параподій.
Рис. 3. Параподія Nereis pelagic;i:
1 - спинний вусик; 2 - нотоподія; 3 - опорні щетинки; 4 - невроподія; 5 - черевний вусик; 6 щетинки невроподи; 7 - щетинки нотогодії
Параподія — нечленистий виріст стінки тіла, що складається з суцільної частини і двох гілок — спинної (нотоподія) та черевної (невроподія). На нотоподії та невроподії розташовані органи хімічного чуття й дотику —відповідно спинний та черевний вусики. У багатьох поліхет спинний вусик перетворюється на зябра. Як правило, усередині параподії є пара товстих опорних щетинок, а назовні стирчать пучечки тонких щетинок досить складної будови, що різняться довжиною, товщиною тощо. Щетинки складаються з хітиноподібної речовини. Параподії рухаються спереду назад: чіпляючись щетинками за субстрат, тварина просувається вперед, синусоїдально звиваючись, при цьому обидві параподії одного сегмента рухаються одночасно в протилежних напрямках. Плаваючі форми мають ластоподібні параподії; у деяких спинна лопать зникає. При плаванні синусоїдальний рух зберігається. У сидячих та риючих форм параподії частково редукуються чи зовсім відсутні, як у Polygordius.
Анальна лопать (пігідій) у багатьох форм, що ведуть активний спосіб життя, має пару органів чуття — анальні вусики.
Зовні тіло поліхет укрите тоненьким шаром пружної кутикули, до складу якої входять колагенові волокна; хітин відсутній. Кутикула виконує тільки захисну функцію. Під кутикулою розташований одношаровий епітелій (епідерміс), де часто знаходяться залозисті клітини (рис. 4). Вони найкраще розвинені у сидячих поліхет. Їхній секрет — слизоподібна речовина — може тверднути, утворюючи навколо тіла захисну хітиноїдну трубку, що часом просякнута вуглекислим кальцієм. У деяких форм частина епітелію війчаста, і війки утворюють кільця та черевну смужку (Protodrilus).
М'язи гладенькі, утворюють два шари: зовнішній кільцевий та внутрішній поздовжній; останній, як правило, не суцільний, а розбитий на дві пари стрічок — спинну та черевну (рис. 5). Кільцевий шар біля параподій розпадається на кілька мускульних пучків, що рухають параподії та щетинки. У кожному сегменті є по парі діагональних м'язів, які навкіс перетинають порожнину тіла, прикріплюючись одним кінцем до базальної мембрани епідермісу по обидва боки черевного нервового ланцюжка, іншим — біля основи параподій. Вони також беруть участь у русі параподій. У товщі м'язів є порожнини, всередині яких містяться опорні щетинки параподій. Вони зв'язані зі стінкою тіла спеціальними м'язами. У перистомії є спеціальні м'язи, що вип'ячують букальний відділ (див. далі), та м'язи, що втягують глотку разом із букальним відділом.
Ізсередини до мускулатури шкірно-м'язового мішка щільно прилягає шар клітин —перигонеальний епітелій, або целотелій, він вистилає вторинну порожнину тіла — целом (див. рис. 1).
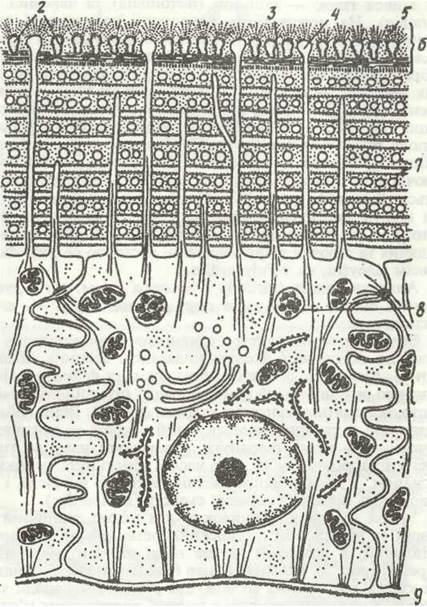
Рис. 4. Схема будови кутикули поліхет за даними електронної мікроскопії:
1 - кутикула; 2 - еліпсоїдні тіла; 3 - пластинчастий шар; 4 - відростки епідермальних клітин; 5 - мукопротеїновий шар; 6— зовнішній шар кутикули без колагенових волокон; 7 - колагенові волокна; 8 - гранули пігменту; 9 - базальна мембрана; 10 - клітина епідермісу
Целом, як уже зазначалося, складається з пари мішків у кожному сегменті. Лівий та правий мітки огортають кишечник і, стикаючись над і під ним, утворюють двошарову поздовжню перетинку — брижу (мезентерій), на якій підвішений кишечник. Стінки сусідніх целомічних мішків утворюють між сегментами двошарові перетинки, так звані септи, або дисепименти. У септах часто є отвори, що з'єднують целоми сусідніх сегментів. У перистомії, як правило, септи відсутні. Целомічна рідина прозора, має амебощні клітини. Целомічні мішки кожного сегмента поліхет є своєрідним гідроскелетом, на який опираються м'язи при русі тварини.
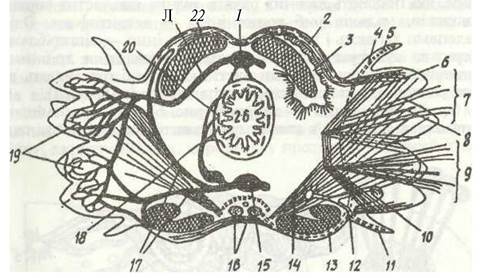
Рис. 5. Схема поперечного зрізу через сегмент тулуба Nereis:
1 - спинна судина; 2 - кільцева мускулатура; 3 - ціліофагоцитарний орган; 4 - спинний вусик параподії; 5 — його нерв; 6 - м'язи щетинок; 7- нотоподія; 8- опорна щетинка; 9 - невроподія; 10 - метанефридій; 11 - черевний вусик; 12 - видільний отвір; 13 - параподіальний нерв; 14 — діагональний м'яз; 15 - черевна судина; 16 - черевний нервовий ланцюжок; 17 - судина, що несе кров від черевної судини до параподії; 18 - внутрішня кільцева судина; 19 - капіляри параподії; 20 — навколокишковий синус; 21 - судина, що несе кров від параподії до спинної судини; 22 - поздовжня мускулатура
Травна система починається ротовим отвором на перистомії, проходить через усе тіло у вигляді прямої трубки, яка складається з букального відділу і глотки, стравоходу, середньої та задньої кишок. На межі букального відділу і глотки у багатьох видів поліхет знаходяться хітинощні щелепи (див. рис. 2). Букальний відділ під час живлення може вивертатися назовні, тоді глотка переміщується вперед, а щелепи опиняються на передньому кінці букального відділу і захоплюють їжу. Глотка мускуляста, з невеликим просвітом. У передню частину стравоходу відкривається пара слинних залоз. Довга середня кишка має складчасті стінки, в її епітелії є багато залозистих клітин, що виділяють травні соки з ферментами; тут їжа перетравлюється і всмоктується. Коротенька задня кишка закінчується анальним отвором на пігідії.
Поліхети живляться по-різному: одні з них — активні хижаки, які поїдають дрібних рачків, різних кільчастих червів, молюсків, мальків риб тощо; інші — детритофаги. Одні щелепами хапають і утримують здобич, інші — зішкрібають детрит із субстрату. Сидячі поліхети живляться дрібними організмами й органічними частинками, які підганяють до рота розвиненими головними щупальцями (пальтами).
Органи виділення — нефридії (рис. 6) завжди розміщені метамерно, попарно в кожному сегменті. Внутрішній кінець кожного нефридія знаходиться в целомічній порожнині сегмента, канал нефридія проходить через дисепимент у наступний сегмент і там відкривається на бічній стороні тіла назовні (див. рис. І). Нефридії мають різну будову. В деяких поліхет є ще протонефридії, які, проте, відрізняються від таких у плоских червів. Замість термінальної клітини на кінці канальців у них є кілька булавоподібних клітин — соленошлів, кожний з яких сліпим кінцем виступає в целом, а в порожнину нефридія відкривається каналом, усередині якого є джгутик. Сукупність джгутиків соленоцитів виконує функцію миготливого полум'я. У багатьох поліхет на місці соленоцитів утворюється лійка з війками, що відкривається в целом. Органи виділення такого типу називаються метанефридіями. Канал нефридія дуже довгий, звивистий, обплетений кровоносними капілярами і оточений компактною залозистою масою, що складається з целомічного епітелію.
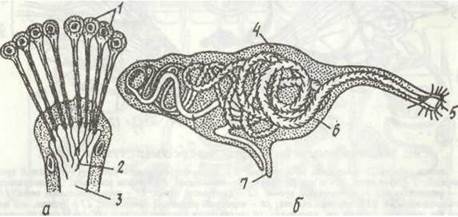
Рис. 6. Органи виділення поліхет:
а — кінцевий відділ нефридія Phyllodoce з соленоцитами; б — метанефридій Nereis; 1 - соленоцити; 2 - миготливе полум'я;
3 - канадець; 4 - залозиста маса; 5 - лійка; б - нефридіальний канал; 7 - видільний отвір
Нефридії виконують функцію осморегуляції та виділення розчинених продуктів обміну.
Дослідження роботи метанефридія показало, що через лійку безперервно надходить целомічна рідина. Рухаючись по каналу, ця рідина зазнає змін. З неї в кров усмоктуються деякі іони, поживні речовини, а з крові до неї надходять продукти дисиміляції, які виводяться назовні у вигляді водного розчину.
Часто з кінцевим відділом нефридія пов'язані так звані целомодукти, що виводять з целому назовні статеві клітини. На відміну від нефридіїв — похідних ектодерми, целомодукти — це органи мезодермального походження. Вони містяться в сегментах попарно. Кожний з них має широку лійку, вкриту війками, коротеньку протоку й вивідний отвір.
Дуже рідко в поліхет є і целомодукти, і нефридії, як, наприклад, у родині Capitellidae (рис. 7). У більшості поліхет нефридії й целомодукти об'єднуються в єдиний орган, що одночасно виконує функції виведення з організму надлишків води, розчинених у ній продуктів метаболізму та статевих клітин. Такі органи звуться нефроміксіями і мають різноманітну будову. В одних випадках, наприклад у Irmalatifrons, лійка целомодукта зростається з нефридіальним каналом і через неї виводяться статеві клітини та екскрети. В інших — на одному нефридіальному каналі сидять лійка целомодукта і кінцева частина протонефридія (соленоцити), тобто вони мають спільний вивідний канал (Alciopacantrainii). Часто кількість органів виділення скорочується (олігомеризація), особливо у сидячих поліхет.
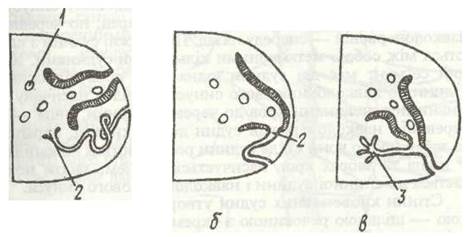
Рис. 7. Схема взаємовідносин між нефридшми та целомодуктами поліхет:
а — самостійні целомодукт і метанефридій; б, в —нефроміксії (стінки нефридіїв зображено чорними, стінки целомодуктів заштриховано); 1 - статеві клітини; 2 - метанефридій; 3 - протонефридш
Крім нефридіїв, видільну функцію виконують спеціальні клітини, що розташовані на стінках кровоносних судин і здатні нагромаджувати продукти азотного обміну (гуанін, солі сечової кислоти тощо). Ці клітини згодом відмирають, їх вміст потрапляє до целома, а звідти через нефридії — назовні. Такі клітини називаються хлорагогетими.
Кровоносна система (див. рис. 5) складається зі спинної та черевної судин, що залягають відповідно у спинному і черевному мезентеріях над і під кишечником. Крім цих судин, навколо кишечника між його зовнішньою стінкою та целомічним епітелієм є вузький щілиноподібний простір — навколокишковий синус. У великих поліхет замість нього утворюється навколокишкове плетиво судин. Ще одна поздовжня судима тягнеться вздовж черевного нервового ланцюжка (навколонервова). По навколокишковому синусу та спинній судині кров рухається ззаду наперед; по черевній і навколонервовій —спереду назад. Поздовжні судини з'єднуються між собою метамерними кільцевими судинами. Кожний сегмент має дві судини: одна несе кров від спинної судини до навколокишкового синуса, а звідти насичену поживними речовинами кров до черевної судини, інша — від черевної і навколонервової судин до стінки тіла, параподій та зябер (якщо вони є), де судини розпадаються на капіляри. У шкірі та зябрах кров насичується киснем, звідти повертається до спинної судини і навколокишкового синуса.
Стінки кровоносних судин утворені опорною пластинкою — щільною речовиною з окремими вкрапленими в неї клітинами. Великі судини (спинна, деякі кільцеві) мають мускулатуру, завдяки якій пульсують. Кров поліхет розносить поживні речовини від кишечника до органів тіла, а також виконує дихальну функцію. У багатьох видів вона містить у розчиненому вигляді дихальні пігменти — гемоглобін чи хлорокруорин.
У частини видів спеціалізовані органи дихання відсутні, і газообмін відбувається через покриви тіла, особливо в параподіях. Багато видів поліхет мають зябра різної будови: у частини сидячих поліхет на зябра перетворюються пальпи або перистомальні вусики; у більшості — спинний вусик параподій. Зябра мають листоподібну, пірчасту або кущоподібну форму. Усередині зябер є кровоносні капіляри, і через їх поверхню відбувається газообмін. Як правило, зябра розташовані не на всіх сегментах, а лише на певних ділянках тіла.
Нервова система (рис. 8) складається з головного мозку — парного надглоткового ганглія, від якого відходять навкологлоткові конекгиви , що огинають глотку і з’єднуються гад нею з парою черевних стовбурів, які тягнуться вздовж усього тіла. У деяких поліхет (Polygordius, Protodrilus) на черевних стовбурах немає гангліїв, уздовж стовбурів розташовані нервові клітини; в інших (Аеlоsoma) — відособлені ганглії ледь помітні; в більшості поліхет у кожному сегменті тіла є пара добре розвинених гангліїв. У примітивних форм (Aelosoma) черевні стовбури широко розставлені, ганглії з'єднані поперечними комісурами, утворюється «нервова драбина».
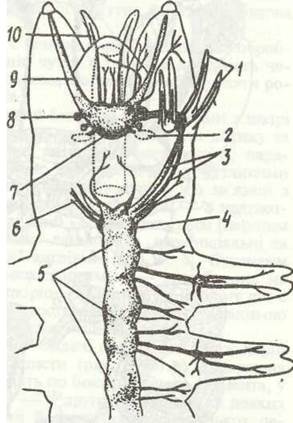
Рис. 8. Нервова система передньої частини тіла Nereis:
1 - перистомальні вусики; 2 ~ нюхальні органи; 3 - навкологлоткові конективи; 4 - підглотковий ганглій; 5 - черевний нервовий ланцюжок; 6 - нерви перистомію; 7 - надглотковий ганглій; 8— очі; 9 - пальпальний нерв; 10 - антена
У більшості видів стовбури зближені, комісури вкорочені, а парні черевні ганглії злиті, в результаті чого утворюється нервовий ланцюжок.
Від кожного ганглія черевного нервового ланцюжка відходять нерви, що іннервують мускулатуру шкірно-м'язового мішка і параподій відповідного сегмента. Кожний ганглій приймає відростки чутливих нейронів, що входять до складу різних рецепторів (наприклад, дотику). Ганглії черевного нервового ланцюжка забезпечують чутливість і рухові реакції в межах відповідного сегмента.
Надглотковий ганглій поліхет —досить складне утворення (рис. 9, а). Він складається з трьох відділів: переднього, середнього й заднього мозку. Передній мозок іннервує пальпи, середній — очі й антени, задній — нюхальні ямки. У бродячих поліхет у середньому мозку є так звані стебельчасті, або грибоподібні, тіла — пара компактних скупчень нервових клітин грибоподібної форми. Стебельчасті тіла є вищим асоціативним центром мозку. У сидячих поліхет, яким не властивий активний пошук їжі, вони відсутні, й загальна будова мозку значно спрощена.
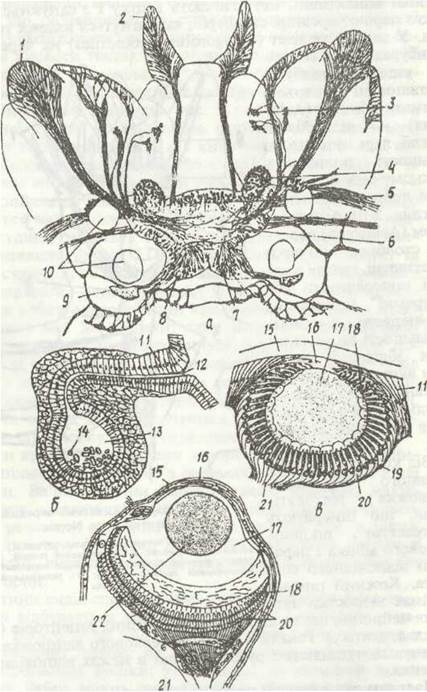
Рис. 9. Органи чуття поліхет:
а- чутливі закінчення на простомії Nereis; б — статоцист Arenicola marina; а, г — перерізи через око
NereisTa Alciopa cantramii відповідно; 1 - пальпи з їхніми чутливими клітинами; 2 - антени; 3 - нервові закінчення на м'язах; 4 -стебельчасті тіла; 5 - передній мозок; 6 - середній мозок; 7 - задній мозок; 8 - татливі клітини нюхальних органів; 9 - нюхальні органи; 10 - очі; 11 - епідерміс; 12 — протока; 13 - його пухирець; 14 - статоліти; 15 - кутикула; 16 рогівка; 17 - склоподібне тіло; 18 - чутливі палички; 19 - пігментні клітини; 20 - зорові клітини сітківки; 21 - нерв; 22 — кришталик
Надглотковий ганглій є центром, який одержує та обробляє інформацію від органів чуття й регулює діяльність черевних нервових стовбурів, координуючи та об'єднуючи роботу сегментарних гангліїв.
Органи чуття представлені багатьма рецепторами в шкірі і, крім того, спеціалізованими органами. Органи дотику та хімічного чуття — пальпи, антени, перистомальні, параподіальні та пігідіальні вусики. Вони густо вкриті чутливими клітинами, відростки яких утворюють нерви, що зв'язані з гангліями: пальпальний та антенальний нерви — з надглотковим ганглієм, перистомальний —з підглотковим (першим ганглієм черевного нервового ланцюжка), параподіальні та анальні — з відповідними гангліями (рис. 9, а). Органами хімічного чуття є також нюхальні ямки — заглиблення на простоміумі, що оточені шкірним валиком; на їхньому дні є чутливі війчасті клітини, відростки яких зв'язані із задньою частиною надглоткового ганглія.
У багатьох видів підкласу Сидячі поліхети є також парні органи рівноваги — статоцисти (рис. 9, б). У піскожила (Arenicola marina) вони лежать по боках першого сегмента, у сабелід (родина Sabeilidae) — у другому сегменті, в деяких видів статоцисти метамерні й зустрічаються в кількох передніх сегментах, наприклад у Scoloplos — у ІV-XІІІ сегментах. Кожен з них має вигляд устеленого чутливими клітинами пухирця, що зв'язаний із зовнішнім середовищем вузьким каналом. Усередині пухирця знаходяться конкреції — «слухові камінці». Статоцисти сприймають коливання води або ґрунту, в результаті чого тварина ховається в трубку. Бродячі поліхети статоцистів не мають.
Майже всі поліхети мають очі різної будови (рис. 9, в, г). Найпростіші вони у бродячих поліхет — у вигляді бокалоподібного заглиблення ектодерми. Епітелій, що його вистилає, відіграє роль сітківки й складається з клітин двох типів: світлочутливих та пігментних. Світлочутливі клітини продовжуються в нервові волокна, що утворюють зоровий нерв, зв'язаний із надглотковим ганглієм. Середина бокала заповнена прозорою драглистою масою — склоподібним тілом. У деяких хижих поліхет (наприклад, у планктонної Аісуора) око ще більш ускладнене: воно відокремлюється від шкіри, перетворюючись на замкнений міхур, де формуються двоопуклий кришталик та склоподібне тіло, а епітелій над кришталиком стає прозорим, утворюючі рогівку. Таке око навіть здатне до акомодації: завдяки дії скоротливих волоконець кришталик може наближатися до сітківки або віддалятись від неї, що дає змогу тварині розглядати предмети на різних віддалях (рис. 9, г). Ці очі належать до типу неінвертованих. У бродячих поліхет вони (два або чотири) розташовані на спинній стороні простоміума.
У сидячих поліхет, які живуть у трубках, численні очі розвиваються на щупальцях, пальпах, параподіях і навіть на пігідії. Вони сприймають лише зміну ступеня освітлення, що спричиняє реакцію ховання в трубку. Ці очі мають таку ж будову, що й у турбелярій, і належать до інвертованого типу.
Поліхети, як правило, роздільностатеві тварини. Статевий диморфізм у них відсутній. Гонади утворюються метамерно в більшій частині або кількох сегментах у стінках целома. Статеві продукти через розриви целомічного епітелію потрапляють до целома, де дозрівають, плаваючи в целомічній рідині. Назовні вони виходять по-різному: через розриви стінок сегментів (Nereis, Eunice тощо), іноді через спеціальні статеві протоки — парні целомодукти. У більшості поліхет лійки целомодуктів сполучаються з нефридіями, утворюючи різноманітні нефроміксії (див. рис. 7). Запліднення зовнішнє.
У багатьох видів гонади розвиваються лише в певній, переважно задній частині тіла, що зветься епітокною. Вона різко відрізняється від передньої атокної частини: на її параподіях з'являються широкі плавальні лопаті й довші щетинки, змінюється колір, редукується кишечник.
Поліхети розмножуються також нестатевим способом. Іноді нестатевий і статевий способи розмноження правильно чергуються, тобто має місце метагенез.
Нестатеве розмноження найчастіше відбувається поперечним поділом тіла. Частини, що відділилися, відновлюють голову або хвостовий відділ. У видів роду Ctenodriius тіло може розпадатися на кілька частин; до складу кожної з них входить від одного до шести сегментів. Кожна частина перетворюється на нового черва. Таке розмноження не пов’язане із статевим дозріванням; досяглий певних розмірів, черв автоматично розпадається на кілька частин. У деяких видів цього роду статеве розмноження взагалі невідоме. Значно рідше відбувається брунькування (наприклад, у Syllis ramosa). На тілі утворюється багато бічних бруньок, з яких розвивається тільки задня частина черва, голова формується вже після відшнурування бруньки (рис. 10, а). Все це свідчить про здатність поліхет до регенерації.
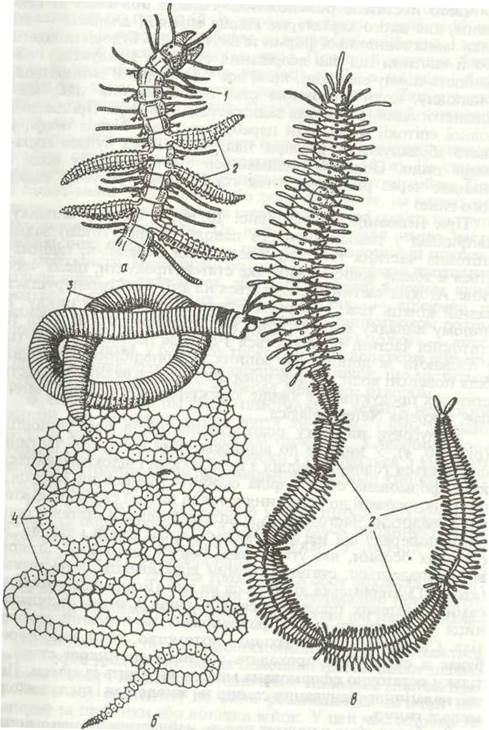
Рис. 10. Розмноження поліхет:
А - бічне брунькування Syllis ramosa; б - неповна епітокія тихоокеанського палоло Eunice viridis; в - нестатеве розмноження Autoiltus; 1 - атокна та 2 - енітокні особини; 3 — атокна частина тіла; 4- епітокна частина тіла
Часто нестатеве розмноження лоліхет пов'язане із статевим, для якого характерне явище епітокії. Епітокія — це різка зміна зовнішньої форми та внутрішньої будови поліхети або її частини під час дозрівання статевих продуктів. Розрізняють повну епітокію, коли все тіло тварини змінюється, і неповну, коли епітокною стає лише частина тіла, здебільшого задня, а передня залишається атокною. Прикладом повної епітокії може бути перебудова тіла у Nereis virens, у якого збільшуються розміри тіла, змінюється будова пара- подій тощо. Поліхета підіймається з дна в товщу води і викидає через розриви стінок тіла статеві продукти, після чого гине.
При неповній епітокії (рис. 10, б), в одному випадку, наприклад у тихоокеанського палоло (Eunice viridis) задня епітокна частина тіла відривається від передньої, підіймається в товщу води, де викидає статеві продукти, після чого гине. Атокна частина залишається на дні, у неї регенерується задній кінець тіла, і тварина готується до нового циклу. В іншому випадку цілі тварини, які складаються з атокної та епітокної частин, підіймаються з дна, де переважно живуть, і плавають за допомогою міцних веслоподібних параподій біля поверхні води; згодом вони випускають велику кількість статевих продуктів. Це явище характерне для родини Nereidae, зокрема Nereis pelagica.
Заслуговує на увагу розмноження видів роду Autolitus (рис. 10, в). У них ще до відокремлення епітокної частини формується головний відділ з придатками і відокремлюється вже сформована статевозріла особина (самець чи самиця), що пристосовані до існування в товщі води й складної статевої поведінки. Часто ще до відділення першої статевої особини попереду від неї утворюється друга, третя і так до 30 статевих особин, які утворюють довгий ланцюжок. Згодом він розпадається, статеві особини відпливають, а нестатева (атокна) материнська лишається на дні. Після виділення біля самиці статевих продуктів самець гине. Самиця відкладає яйця у виводкову сумку, розташовану на черевній стороні тіла. Своїм тілом вона захищає потомство, доки воно перебуває в сумці. Тут проходять усі його личинкові стадії, і тільки остаточно сформована молодь виходить із сумки. Під час пелагічного існування самиці не живляться і після виходу молоді гинуть.
Дробіння яйця у поліхет повне, найчастіше нерівномірне, спіральне і детерміноване, тобто вже на ранніх стадіях точно визначено подальший розвиток кожного бластомера. В результаті дробіння утворюється кулеподібна бластула. Гаструляція відбувається шляхом інвагінації або епіболії. Гаструла має бластопор на вегетативному полюсі. Потім бластопор видовжується по одній із сторін гаструли (майбутній черевній) до її екватора, набуває щілиноподібної форми і заростає по всій довжині, ззаду наперед. Передній залишок бластопора перетворюється на ротовий отвір, а на місці його заднього краю проривається анальний отвір. Так утворюється наскрізний кишечник. Подібний розвиток й у нематод (див. кн.1, с. 294).
Прямий розвиток у поліхет буває дуже рідко; як правило, з яйця виходить личинка трохофора (рис. 11, а). Це типова планктонна личинка, що рухається за допомогою війок. Тіло її більш-менш кулеподібне або дещо витягнуте, на верхньому полюсі розташована тім'яна китиця війок, які сидять на групі ектодермальних клітин. Це орган чуття, біля нього часто лежать інші органи чуття — пара вічок, коротенькі щупальця, статоцисти. Приблизно по екватор/трохофори проходить кільце війок — прототрох, або передротовий віночок, який поділяє тіло личинки на верхню півкулю — еггіеферу та нижню — гіпосферу. Безпосередньо під прототрохом розташований рот, на нижньому (вегетативному) полюсі — анус. Крім прототроха, у багатьох τροχοφορ є ще додаткові війчасті кільця — метатрох, або післяротовий віночок, що міститься позаду рота, та телотрох — перед анальним отвором.
В епісфері, під тім'яною пластинкою, лежить нервовий ганглій, що іннервує органи чуття личинки. З ним зв'язані нервові меридіональні і кільцеві стовбури. Кишечник трохофори має вигляд вигнутої трубки і складається з ектодермального стравоходу, ентодермальної середньої кишки та ектодермальної задньої.
Між стінкою тіла трохофори і кишечником розташована первинна порожнина тіла, в якій містяться поодинокі мезодермальні клітини (мезенхіма) та м'язи. У нижній півкулі личинки є пара протонефридіїв.
Біля нижнього полюса трохофори з обох боків від кишечника лежать дві великі клітини — мезодермальні телобласти, іноді вони починають ділитися, утворюючи дві мезодермальні смужки.
Після деякого періоду планктонного життя починається метаморфоз трохофори. Задня половина тіла личинки значно подовжується і поділяється одночасно на кілька (найчастіше 3, 7 або 13) сегментів, на яких розвиваються зачатки па- раподій та щетинки або віночки війок. У цей час телобласіи починають ділитись, утворюючи два тяжі — мезодермальні смужки, що лежать по боках від кишечника. Якщо ці смужки вже були у трохофори, то тепер їх клітини посилено діляться. Згодом усередині кожної з мезодермальних смужок клітини розсуваються, й утворюється порожнина — зачаток целома.
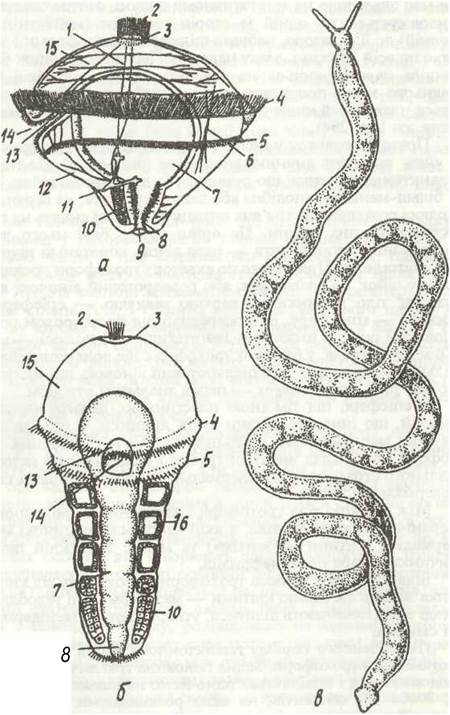
Рис.11. Метаморфоз Polygordius:
а - трохофора; б -її дальший розвиток; в -доросла особина; 1 - нервові стовбури; 2-тім'яна китиця; 3 - тім'яна пластинка; 4 - прототрох; 5 - метатрох; 6 - м'язи; 7 - середня кишка; 8 - задня кишка; 9 - анус; 10 - мезодермальна смужка; 11 - протонефридій; 12 - гіпосфера; 13 - передня кишка; 14 - рот, 15 - епісфера; 16 - целомічні мішки
Найчастіше целомічні мішки залишаються у личинки суцільними, проте в деяких видів вони розділяються, утворюючи по парі целомів у кожному сегменті. Личинку, що має кілька сегментів, називають метатрохофорою. її тіло складається з головної лопаті (простомія), яка становить майже незмінену верхню півкулю трохофори, кількох сегментів і маленької анальної лопаті (пігідія) — ділянки нижньої півкулі трохофори навколо ануса (рис. 12). Сегменти, що утворились у метатрохофори, називають личинковими, або ларвальними.
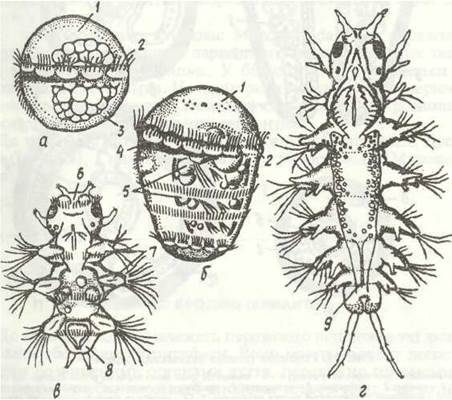
Рис. 12. Личинкові стадії Nereis:
а -трохофора; б - метатрохофора; в - нектохета; г - молода донна форма; 1 -епісфера; 2 - прототрох; 3 - гіпосфера; 4 - рот; 5 - зачатки параподій; 6 - простомій; 7 - параподії; 8 - піддій; 9 - зона росту
У багатьох видів поліхет метатрохофора перетворюється на наступну личинкову стадію — нектохету, в якої вже з'являються головні придатки — антени, пальпи, лопаті параподій із щетинками, тобто вона більше схожа на дорослу особину, але має лите ларвальні сегменти і веде планктонний спосіб життя (рис. 12, в). На епісфері в неї розвиваються очі, антени, органи нюху. Зв'язані з ними нервові клітини, об'єднуючись, утворюють головний мозок (надглотковий ганглій). В ектодермі черевної сторони тіла з'являються зачатки гангліїв черевного нервового ланцюжка. Згодом на задньому кінці личинки, перед анальним отвором, утворюється зона росту, спереду від якої один за одним формуються сегменти, що звуться постларва (рис. 12, г).
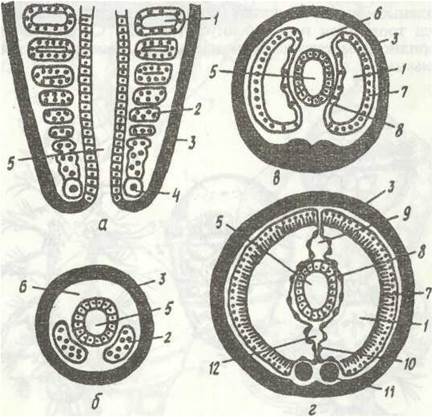
Рис. 13. Розвиток целома кільчастих червів:
а - фронтальний зріз через зону росту; б - г — поперечні зрізи через черва на різній відстані від зони росту; 1 - целом; 2 - мезодерма; 3 — ектодерма; 4 - телобласт; 5 - кишка; б - первинна порожнина тіла; 1,8- зовнішній та внутрішній листки мезодерми; 9, 12 - спинна 1-а черевна кровоносні судини; 10 - мезентерій; 11 - черевний нервовий ланцюжок
У зоні росту клітини мезодермальних смужок інтенсивно діляться, від них відділяються парні зачатки целомічних мішків. Кожен сегмент, що утворюється, має пару таких зачатків, які, розростаючись, утворюють целомічні мішки (рис. 13). Зовнішньою стороною кожен мішок підстилає шкірно-м'язовий мішок відповідного сегмента, внутрішньою оточує кишечник. Стінками целомічних мішків, що прилягають один до одного над і під кишечником, утворюються спинна і черевна брижі, а в місці стикання розташованих один за одним мішків — поперечні перетинки між сегментами — септи, або дисепименти. Кровоносні судини виникають між
двома шарами целомічного епітелію у спинній та черевній брижах (мезентеріях). За рахунок клітин мезодермальних смужок формуються також мускулатура шкірно-м'язового мішка і кишечника, клітини целомічної рідини, крові, хлорагогенні клітини. Утворення сегментів у зоні росту триває дуже довго, іноді до кінця життя черва, що призводить до значного збільшення їх кількості.
Отже, у поліхет є дві групи сегментів, що різняться способом утворення. Ларвальні сегменти виникають першими, всі одночасно, розчленуванням нижньої півкулі трохофори. Постларвальні сегменти утворюються послідовно один за одним у зоні росту.
У частини поліхет (підклас Myzostomida) тіло протягом життя складається лише з ларвальних сегментів. Таких тварин називають олігомерними. У більшості ж формуються і постларвальні сегменти. Це полімерні кільчаки. У полімерних кільчастих червів гонади, целомодукти та нефридії розвиваються лише в постларвальних сегментах.
До класу Багатощетинкових належать три підкласи: Бродячі (Errantia), Сидячі (Sedentaria) та Мізостоміди (Myzostomida).