ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 3 - Г.Й. Щербак - 1997
ТИП ПОГОНОФОРИ (POGONOPHORA)
КЛАС БЕЗВУЗДЕЧКОВІ (AFRENULATA, АБО VESTIМENTIFERA)
Безвуздечкові — досить великі погонофори, завдовжки до 1,5 м при діаметрі 1 см, що живуть на дні морів і океанів поблизу гідротермальних джерел та холодних виходів води, насиченої сірководнем або метаном (рис. 157). Перший вид — Lamellibrachia barhami — було описано в 1969 р. американським вченим Веббом, для якого він встановив новий клас Afrenulata, віднісши всіх інших відомих на той час погонофор до класу Frenulata. Поки що відомо 15 видів вестиментифер.
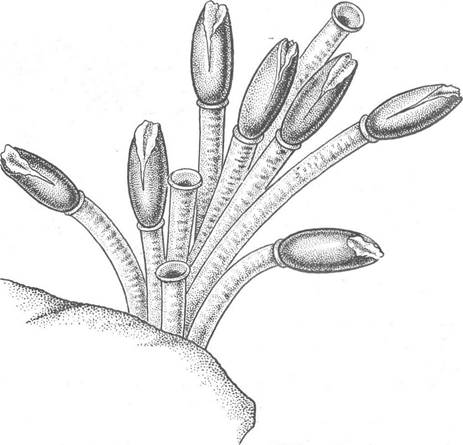
Рис. 157. Угруповання особин Riftia pachyptila
Тіло безвуздечкових, як і інших погонофор, складається з чотирьох відділів (рис. 158). На передньому з них міститься пара мускулястих виростів, направлених вперед і з’єднаних між собою кутикулярними перетинками, що утворюють так званий абтуракул (від латинського abturo — затикати). На передньому кінці абтуракула розташована особлива кутикулярна кришечка, що закриває вхід у трубку, коли тварина ховається в ній. Обтуракул розвивається з личинкових щупалець, він має целомічний канал з кровоносними судинами, які підтримуються мезентерієм і іннервуються тим самим нервом, що й щупальця. Відрізняються абтуракули від щупалець більшим розвитком сполучної тканини, відсутністю пінул та війчастих клітин епідермісу. По боках від абтуракула розташовані численні (у Riftia pachyptila до 300) щупальцеві, або, як їх часто називають, зяброві пластинки, які складаються з сотень щупалець (зябрових ниток), щільно з’єднаних між собою кутикулою. Вільними лишаються лише їх передні кінці, на яких містяться пінули, які в безвуздечкових є вип’ячуваннями стінки щупальця, наповненими кров’ю, що над-

Рис. 158. Схема будови тіла вестиментифери:
1 — війчаста смужка; 2 — щупальцеві пластинки; 3 — абтуракул; 4 — вестимент; 5, 6 — тулубний та хвостовий відділи
ходить сюди з кровоносних судин зябрових ниток. Щупальцева корона має яскраво-червоний колір через кров, що просвічує крізь покриви.
Другий відділ має два крилоподібних вирости тіла з сильно розвиненою мускулатурою, що загортаються на спинну сторону тварини, яка неначе загортає на собі покривало, тому цей відділ називають вестиментом (від лат. vestimentum — одяг, покривало). На черевній стороні другого відділу є широка війчаста смужка. Вуздечки немає.
Третій, тулубний, відділ, який становить половину, а у великих Riftia до 80 % довжини тіла, не поділений на дві частини, як у вуздечкових. Як і зовнішня сторона крилоподібних виростів другого відділу, його покриви вкриті бородавчастими залозами, які, можливо, сек- ретують речовину трубки.
Четвертий, хвостовий, відділ, завдовжки 2—3 см складається з кількох десятків сегментів, відділених один від одного перетинками. Кожний із сегментів має поясок з дрібних зубчастих щетинок.
Основну частину тулубного відділу займає трофосома — живляча залоза, яка складається з численних трубочок, обплетених кровоносними судинами і повністю заповнених симбіотичними аутотрофними бактеріями, які окислюють сірководень або метан. Енергія, що при цьому звільнюється, використовується в процесі хемосинтезу органічних речовин із вуглекислоти при наявності високоактивних ферментів. Кров постійно приносить бактеріальним симбіонтам кисень та сірководень (H2S), і в кров надходять синтезовані бактеріями органічні речовини, які потрібні для живлення тканин і органів вестиментифери. Дослідження дрібних погонофор із класу Frenulata, зокрема родів Siboglinum та Oligobracliia, що живуть на звичайному морському дні, показали, що і в них у постанулярній частині тулуба є симбіотичні бактерії, які заповнюють клітини целомічного епітелію між кровоносними судинами. Значну частину поживних речовин ці тварини також дістають за рахунок хемосинтезу симбіонітів.
Ембріональний розвиток безвуздечкових не вивчено, відомо лише, що яйця в трубку не відкладаються. Спостереження за Riftia pachyptila в акваріумі показали, шо яйця в неї дрібні, але багаті на жовток. Яйця легші за воду і тому повільно спливають на її поверхню. Це дає змогу припустити, що яйця та личинки безвуздечкових можуть розноситись течіями, що має важливе значення для тварин у разі виснаження термального джерела, біля якого вони живуть, а це відбувається не більше, ніж через 10 років. Молода особина (Ridgeia sp., Oasisia alvinae, Riflia pachyptila), яка тільки що осіла на дно, завдовжки 0,15 мм, відразу починає виділяти трубку, що спочатку має колбоподібну форму, а потім витягується. При довжині 0,25—0,30 мм вестиментифера має пару добре розвинених щупалець (рис. 159), а при довжині 5—6 мм вона вже може втягуватись у трубку і закривати її отвір кришечкою.
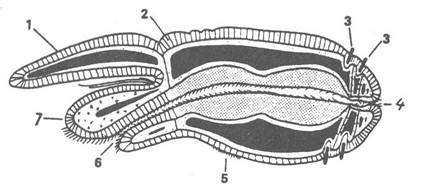
Рис. 159. Схема будови молодої вестиментифери на стадії двох щупалець:
1 — щупальце; 2 — епідерміс; 3 — щетинки; 4 — анус; 5 — черевна війчаста смужка; 6 — рот; 7 — головний виріст
На ранніх стадіях розвитку вестиментифери мають травну систему: рот веде в лійку-глотку, за нею йде війчастий стравохід, який проходить крізь мозок і продовжується в середню кишку; остання переходить в об’ємну задню кишку. Саме середня кишка перетворюється на трофосому. Молоді особини поглинають через рот вільноживучих сірчаних бактерій, які в масі живуть біля сірчаних джерел. При попаданні в середню кишку їх поглинають фагоцити, але не перетравлюють їх, а переносять у складки кишечника, де фагоцити осідають, а сірчані бактерії, що стають симбіонтами, починають розмножуватись. Травна система функціонує ще досить довго —до формування кількох сотень щупалець, приносячи бактеріям кисень та сірководень або метан. Потім функція постачання переходить до кровоносної системи, а кишечник поступово зникає. Спочатку середня кишка відділяється від задньої, яка редукується і зникає разом із анусом. Пізніше втрачається зв’язок зі стравоходом, який деградує, але зберігається в дорослих особин у вигляді тонкого тяжа, що проходить крізь мозок. Вивчення вестиментифер тільки починається.