Основи біоорганічної хімії (навчальний посібник) - Г. О. Сирова - 2018
Тема
Структурна організація білків. Фізико-хімічні властивості білків.
Реакції осадження білків. Денатурація
Актуальність теми. Білки - це особливий клас речовин, які зустрічаються у всіх живих організмах. Вони відіграють важливу роль у життєдіяльності організму. З огляду на структуру білками називаються високомолекулярні полімери, мономерами яких є амінокислоти. У білках знаходяться 20 різних амінокислот. Білки виконують цілий ряд біологічних функцій. Так, білки-ферменти обумовлюють обмін речовин в організмі, є в організмі транспортні білки, які переміщують речовини в крові, лімфі; частина гормонів - це білки або поліпептиди. Процес передачі спадкової інформації не може відбуватися без білків. Усі форми механічного руху обумовлені білками м'язів. Серед функцій білків найважливішими є здатність білків підтримувати онкотичний тиск крові, регулювати обмін води, підтримувати сталість рН, служити джерелом енергії (до 10 % енергії). Без знань про будову і властивості ВМС неможливо пізнати сутність біологічних процесів.
Загальна мета: уміти аналізувати відповідність структури біоорганічних сполук фізіологічним функціям, які вони виконують в організмі, інтерпретувати особливості будови і перетворення в організмі біоорганічних сполук.
Конкретні цілі:
1. Інтерпретувати особливості будови а-амінокислот як основи біополімерів білків, які є структурними компонентами всіх тканин організму.
2. Пояснювати залежність фізико-хімічних властивостей білків від їх амінокислотного складу.
3. Уміти аналізувати якісні реакції на а-амінокислоти для визначення амінокислотного складу білків і використання біуретової реакції для кількісного визначення білків.
Теоретичні питання
1. Білки як біополімери. Способи сполучення а-амінокислот у молекулах білків.
2. Хімічні зв'язки, що формують первинну, вторинну, третинну і четвертинну структуру білка.
3. Глобулярні і фібрилярні білки. Вивчення рівнів структурної організації білкових молекул.
4. Хімічний синтез пептидів і білків.
5. Чинники стабільності існування білків у колоїдних розчинах.
6. Механізм осадження білків. Види осадження білків. Реагенти, що сприяють висолюванню. Необоротне осадження. Чинники, що викликають необоротне осадження.
7. Денатурація, її ознаки. Чинники, що сприяють денатурації білків. Ренатурація.
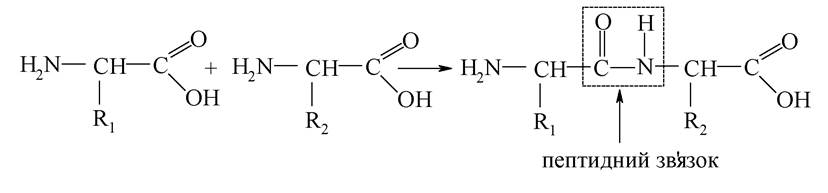
1. Білки - це високомолекулярні нітрогеновмісні сполуки, які мають пептидні (амідні) зв'язки і під час гідролізу розщеплюються на амінокислоти.
Білки також можна розглядати як упорядковані біополімери, що являють собою поліконденсати α-амінокислот. Пептидний зв'язок утворюється в результаті взаємодії карбоксильної групи однієї α-амінокислоти з аміногрупою іншої амінокислоти. Утворені сполуки називаються пептидами або поліпептидами. Унаслідок того, що атом карбону пептидного зв'язку знаходиться в sр2-гібридизації, атоми, які беруть участь в утворенні зв'язку, розташовані в одній площині.
Структура поліпептиду (поліпептидного ланцюга) є основою будови молекули білка. Утворення поліпептидів і білків із залишків амінокислот пов'язане з відщепленням молекул води, тому білки і поліпептиди є поліконденсатами амінокислот:
За кількістю α-амінокислотних залишків, які беруть участь у будові пептиду, розрізняють олігопептиди (низькомолекулярні пептиди), що містять у ланцюзі не більше 10 амінокислотних залишків, і поліпептиди, до складу ланцюга яких входять до 100 амінокислотних залишків.
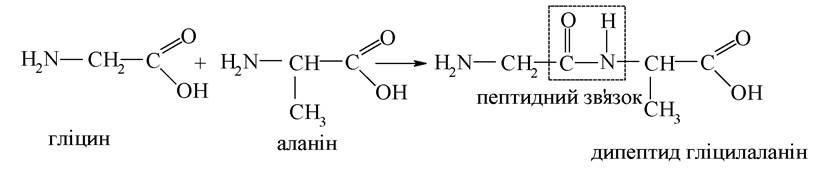
Конструкція поліамідного ланцюга однакова для всього різноманіття пептидів і білків. Цей ланцюг складається з амідних і метинових груп (СН), які чергуються. На одному кінці знаходиться а-амінокислота з вільною NH2- групою. Цей кінець ланцюга називають N-кінцем. На іншому кінці ланцюга є вільна COOH-група, називають цей кінець С-кінцем. Пептидні і білкові ланцюги прийнято записувати, починаючи з N-кінця; радикали а-амінокислот розміщують у бічному ланцюзі. Назву пептидів утворюють від назви відповідних амінокислот. При цьому амінокислоти, які беруть участь в утворенні пептидного ланцюга за рахунок карбоксильної групи, мають закінчення -іл, а ті, які беруть участь як аміногрупа - закінчення -ін.
2. Молекули білків можуть складатися з одного або декількох поліпептидних ланцюгів, кожен з яких має специфічне для цього білка чергування певних а-амінокислотних залишків. Фізико-хімічні і біологічні властивості білків обумовлюються не тільки хімічною будовою поліпептидного ланцюга, але і просторовою конфігурацією всієї молекули. У білках розрізняють чотири рівні структури.
Первинна структура. Амінокислотна послідовність, тобто порядок чергування α-амінокислотних залишків, становить первинну структуру пептидів і білків. У ланцюзі а-амінокислотні залишки поєднані, як зазначалося вище, між собою пептидним (ковалентним) зв'язком. Усі атоми, які входять у пептидну групу, лежать в одній площині:
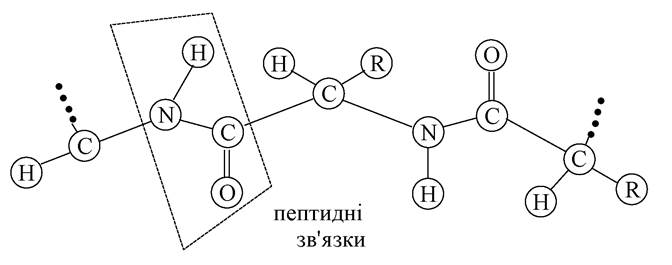
В основі поліпептидного ланцюга чергуються плоскі пептидні групи з одними (СН-R), які здатні обертатися навколо зв'язків.
Пептидні зв'язки впливають не тільки на форму первинної структури, але на більш високі рівні організації молекули білка. Первинна структура - найпростіший рівень структурної організації молекули білка.
Вторинна структура - це просторова конформація поліпептидного ланцюга. Первинна структура білка фіксується в просторі за допомогою водневих зв'язків, що утворюються в основному між атомами оксигену карбонільних груп і атомом гідрогену амідних груп одного і того ж пептидного ланцюга або між різними поліпептидними ланцюгами. Ці зв'язки стабілізують вторинну структуру білкової молекули. Найбільш типовою для білків є а-спіраль, існування якої довели вперше Л.Полінг і Р.Корі. Закручування поліпептидного ланцюга в спіраль відбувається за годинниковою стрілкою. Водневі зв'язки виникають між пептидними групами кожного першого і четвертого, другого і п'ятого та ін. задишка амінокислоти. На кожен виток спіралі доводиться 3,6 амінокислотних залишків. α-Спіральна конформація характерна для глобулярних білків. За наявності SH-груп між ними виникають дисульфідні зв'язки в межах одного поліпептидного ланцюга (рис. 5):
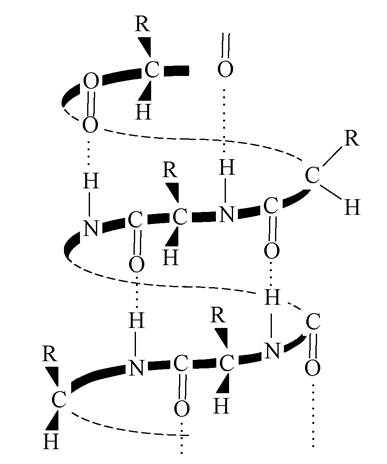
Рис. 5 - α-спіраль
Іншим видом вторинної структури є так звана структура складчастого листка, або β-конформація (рис. 6).
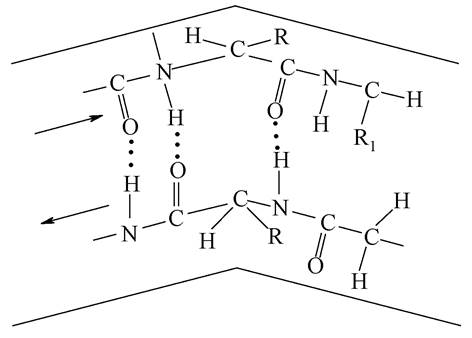
Рис. 6
Вона виникає між антипаралельними ланцюгами, які забезпечують сприятливі умови для утворення водневих зв'язків між ними. Наявність спіральних структур доведено для таких білків, як кератин - білок сполучної тканини, міозин - білок м'язів. Р-конформація знайдена в β-кератині та фіброїні шовку. β-конформація властива фібрилярним білкам.
Під третинною структурою розуміють форму укладання (упаковки) спіралей, тобто вторинних структур з утворенням глобул різної форми.
Тим часом як вторинна структура білка визначається водневими зв'язками, численні вигини поліпептидного ланцюга, що надають білкам третинну структуру, залежать не тільки від пептидних і водневих зв'язків, а і від інших типів взаємодії, а саме сульфідних зв'язків у цистеїні (сульфідний зв'язок), іонної та гідрофобної. Дисульфідний зв'язок належить до нековалентних зв'язків та виникає в результаті відщеплення двох атомів гідрогену від залишків цистеїну. Цими зв'язками можуть з'єднуватися залишки амінокислот в одному поліпептидному ланцюзі, особливо тоді, коли вона утворює певні вигини та спіралі. Також дисульфідні зв'язки можуть утворюватися між залишками цистеїну двох поліпептидних ланцюгів. Стійкість деяких білків (колагену) значною мірою обумовлена «прошиванням» дисульфідними зв'язками витків спіралі, утвореної пептидним ланцюгом молекули білка.
Схема утворення дисульфідного зв'язку така:
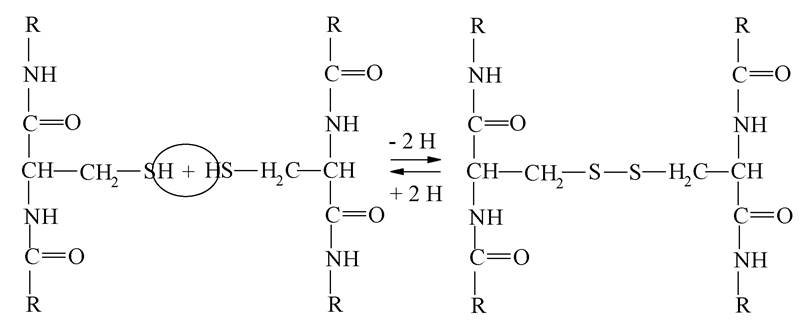
Прикладом білкових структур, що містять дисульфідні зв'язки, можуть бути інсулін та рибонуклеаза.
Іонний зв'язок. Аніонні групи (-СОО-) пептидного ланцюга із залишків аспарагінової або глутамінової кислот та катіонні групи (MN3) залишку лізину як протилежно заряджені можуть у разі зближення притягатися та утворювати сольові зв'язки (містки). Схема сольового зв'язку між двома пептидними ланцюгами:
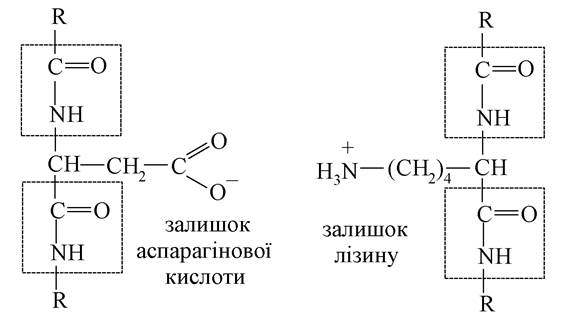
Вони розміщуються на зовнішній поверхні молекули білка та знаходяться в гідратованому стані.
До нековалентних зв'язків належать псевдопептидні зв'язки, що виникають між аміногрупами бічних радикалів лізину, аргініну і СООН- групами бічних радикалів аспарагінової та глутамінової кислот. Звідси і назва цього типу зв'язку - подібний пептидному.
Гідрофобний зв'язок утворюється в результаті зближення неполярних вуглеводневих радикалів таких амінокислот, як валін, лейцин та ін.
Зближення цих радикалів обумовлено характером взаємодії гідрофобних груп поліпептидного ланцюга (метильного -СН3, етильного -С2Н5 та ін.) з водою. Поблизу цих вуглеводневих радикалів молекули води перебудовуються в просторі, утворюючи структуру, що нагадує кристали льоду. У результаті цього гідрофобні групи переміщаються у внутрішні частини молекул, а на поверхні білка розміщуються гідрофільні (-ОН, -СООН, -NH2, =NH, -SH), що і обумовлює гідратацію білків. За умови такого згортання поліпептидних ланцюгів у воді гідрофобні групи білка розміщуються поруч одна з одною, а не з водою.
Схема утворення гідрофобного зв'язку між двома пептидними ланцюгами:
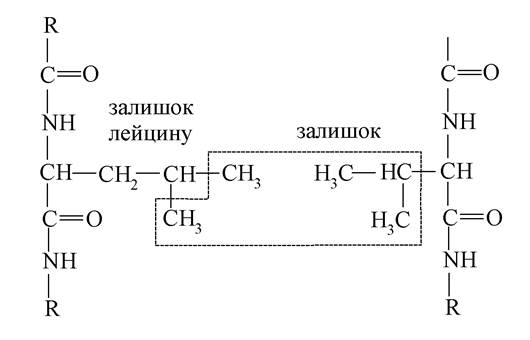
Також у третинній структурі молекул білка мають місце водневі зв'язки. Вони виникають між бічними радикалами амінокислот різних ділянок ланцюга.
Четвертинна структура. Кілька окремих поліпептидних ланцюгів здатні до більш складних утворень - комплексів або агрегатів. При цьому кожен ланцюг, зберігаючи характерну для нього первинну, вторинну та третинну структури, виступає в ролі субодиниці комплексу з більш високим рівнем просторової організації. Такий комплекс являє собою єдине ціле та виконує біологічну функцію, не властиву окремо взятим субодиницям. Четвертинна структура закріплюється за рахунок водневих зв'язків, іонних та гідрофобних взаємодій між субодиничними поліпептидними ланцюгами. Четвертинна структура характерна лише для деяких білків, наприклад гемоглобіну.
У таблиці подано узагальнені дані про зв'язки, які стабілізують різні рівні організації білкової молекули.
Вид структури білка |
Зв'язки, що стабілізують структуру |
Первинна структура (лінійний поліпептидний ланцюг) |
Пептидні зв'язки - між α-аміно- і α-карбоксильними групами амінокислот |
Вторинна структура (α-спіраль, β-структура) |
Водневі зв'язки - між пептидними групами (кожна перша та четверта) одного поліпептидного ланцюга або між пептидними групами суміжних поліпептидних ланцюгів; дисульфідні зв'язки - між SH-групами в межах одного поліпептидного ланцюга |
Третинна структура (глобулярна, фібрилярна) |
Дисульфідні зв'язки - між бічними радикалами амінокислот різних ділянок пептидного ланцюга; водневі зв'язки - між бічними радикалами амінокислот різних ділянок ланцюга; іонні зв'язки - між протилежно зарядженими групами амінокислот кожної субодиниці; гідрофобні взаємодії - між неполярними радикалами амінокислот у водному середовищі |
Четвертинна структура (глобулярна) |
Іонні зв'язки - між протилежно зарядженими групами амінокислот кожної субодиниці; водневі зв'язки - між бічними радикалами амінокислотних залишків кожної субодиниці; гідрофобні взаємодії - між неполярними радикалами амінокислот у водному середовищі |
3. Відома велика кількість білків різних за фізико-хімічними властивостями і біологічною роллю. Для систематизації та вивчення запропоновано кілька класифікацій.
В основі класифікації лежить кілька ознак:
1) форма білка, тобто просторова структура (конформація);
2) фізико-хімічні властивості;
3) хімічний склад білків.
За формою молекул білки класифікують як глобулярні та фібрилярні.
Глобулярні білки. Вони мають форму молекул, подібну до кулястої (від лат. globulus - кулька). До них належить більшість розчинних білків, таких як альбуміни, білки молока, яєць та ін.
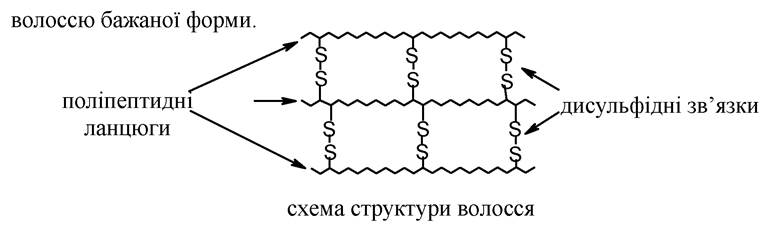
Фібрилярні білки. Вони мають ниткоподібну, витягнуту або фібрилярну форму молекули. Довжина таких молекул у багато разів перевищує їх діаметр. До них належить білок волосся - кератин, білок шовку - фіброїн, білок м'язів - міозин та низка ін. Ці білки за рахунок зазначеної форми можуть стискатися та розпрямлятися, що обумовлює еластичність білків сполучної тканини і скорочення м'язів.
Так, наприклад, волосся складається з фібрилярного білка (кератину). Він містить велику кількість такої амінокислоти, як цистеїн. За рахунок окиснення цистеїнових залишків утворюються дисульфідні містки, що з'єднують поліпептидні ланцюги. Ці містки надають своєрідну форму волоссю людини (кучеряві, прямі). Використовуючи відновники можна зруйнувати дисульфідні містки та надати волоссю іншу форму, а потім подіяти на волосся окиснювальним агентом. Утворюються нові дисульфідні зв'язки, які нададуть
За складом білки поділяються на прості (некон'юговані) і складні (кон'юговані).
Прості білки під час гідролізу розщеплюються на амінокислоти; складні білки, крім амінокислот, у своєму складі містять ще і речовини небілкового характеру, які визначають характер структури і біологічну дію білка, наприклад, гем у гемоглобіні або нуклеїнові кислоти в нуклепротеїнах.
Однією з найважливіших задач сучасної науки є вивчення будови природних поліпептидів, які являють собою складові частини білка. Як зазначалося вище, специфічність білків визначаться двома найважливішими характеристиками - амінокислотним складом і амінокислотною послідовністю.
Амінокислотний склад білка встановлюється таким чином: проводиться повний гідроліз, а потім ідентифікація кислот, які утворилися,
хроматографічними методами. На сьогодні такий аналіз здійснюється автоматично за допомогою амінокислотних аналізаторів.
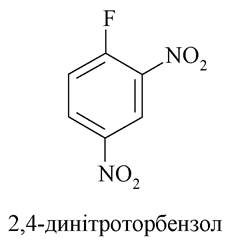
Визначення амінокислотної послідовності проводиться шляхом відщеплення а-амінокислоти з N-кінця або С-кінця з подальшою ідентифікацією відщепленої а-амінокислоти. Досить добре розроблено хімічні способи відщеплення а-амінокислот з N-кінця. Уперше хімічний спосіб запропонував Ф. Сенджер. Пептид обробляють в слабколужному середовищі
2,4-динітрофторбензолом, який взаємодіє з а-амінокислотою на N-кінці, а потім повністю гідролізують отриману сполуку, з гідролізату виділяють і ідентифікують ДНФ-похідне а-амінокислоти, що знаходиться на N-кінці вихідної пептидної молекули.
Інший метод встановлення амінокислотної послідовності називається методом деградації за Едманом. Сутність цього методу подібна першому. Як реагент, що відщеплює а-амінокислоту, використовують фенілізотіоціанат. Перевага цього методу полягає в тому, що під час виділення похідної N-кінцевої амінокислоти інша частина пептидної молекули не руйнується, отже, операцію з відщеплення а-амінокислот можна продовжувати. Цей метод відтворено в автоматичному приладі - секвенаторі, за допомогою якого можна здійснити 40-50 відщеплень.
Для визначення амінокислотної послідовності використовують також частковий кислотний або ферментативний гідроліз. При цьому поліпептид розщеплюється на олігопептиди, які можна розділити та ідентифікувати хроматографічним методом. Часто поліпептид розщеплюють поступово за допомогою специфічних ферментів, здатних розривати пептидні зв'язки.
Структурний аналіз навіть простого поліпептиду, яким би методом він не проводився, вимагає багато часу і сил. Так, наприклад, Ф. Сенджер 10 років займався розшифровкою будови інсуліну, порівняно нескладного поліпептиду, у складі якого міститься 51 а-амінокислотний залишок.
Для визначення в білках а-спіралей і P-структур використовують метод спектрополяриметрії. Метод базується на здатності амінокислотних залишків поліпептидного ланцюга повертати площу поляризованого променя. α-Спіралі і β-структури повертають площу поляризації світла вправо. Між кутом обертання та вмістом у білку α- і β-структур існує пряма залежність. Визначають ступінь спіралізації білків (вміст α-спіралей) за допомогою УФ-спектрофотометрії.
4. Важливим завданням сучасної науки є синтез пептидів заданої будови та фізико-хімічних властивостей. Пептиди заданої будови не вдається отримати прямою конденсацією а-амінокислот, оскільки останні є біфункціональними сполуками. Навіть у найпростішому випадку поєднання двох компонентів, наприклад гліцину і аланіну, утворюється чотири дипептиди. При збільшенні числа амінокислотних залишків кількість потенційних пептидів збільшується. Так, з 20 амінокислотних залишків можливо отримати 1014 пептидів, тобто дуже велику кількість сполук. Цим і обумовлено велике різноманіття білків та пептидів.
Зараз розроблено методику синтезу пептидів, згідно з якою на певних етапах здійснюється захист (блокування) одних функціональних груп і активація інших функціональних груп.
Активованими повинні бути карбоксильна група однієї амінокислоти та аміногрупи іншої амінокислоти, тобто ті функціональні групи, які утворюють пептидний зв'язок. Одночасно блокують аміногрупу першої кислоти і карбоксильну групу іншої амінокислоти.
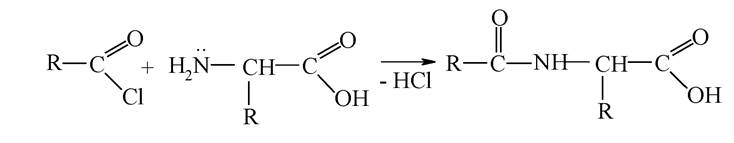
Захист аміногрупи здійснюється ацилюванням:
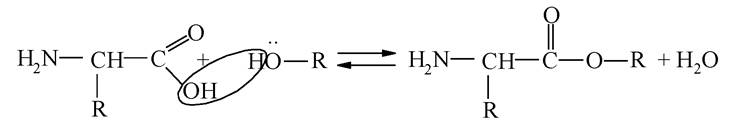
Карбоксильну групу захищають шляхом перетворення її на естер:
Активація карбоксильної групи здійснюється шляхом перетворення її на хлорангідрид:
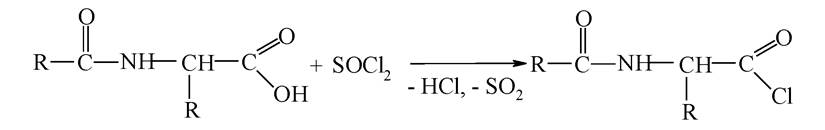
Утворення пептидного зв'язку відбувається за схемою:
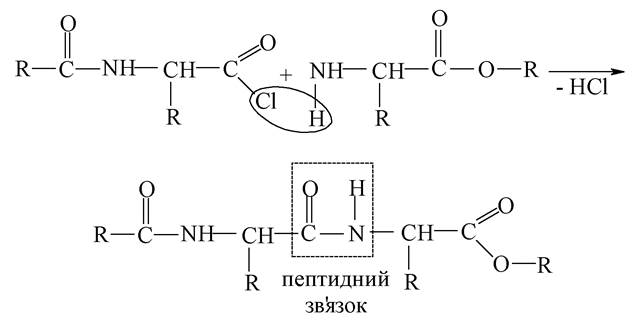
Таким чином, «крок за кроком» послідовно відбувається нарощування ланцюга. При цьому доводиться проводити велику кількість експериментальних операцій з виділення і очищення продукту на кожній стадії, що пов'язане з втратами - вихід продукту не перевищує 1 %.
Більш перспективним методом хімічного синтезу білків є твердофазний синтез, запропонований Б. Мерріфілдом (1963 р.). Цей метод відрізняється від вищенаведеного класичного методу синтезу тим, що використовується твердий носій у вигляді зерен полімерної смоли, поверхня яких містить хлорметильні групи -CH2Cl (якірні групи). На початковій стадії перша а-амінокислота із захищеною МН2-групою прикріплюється (сідає на якір) до поверхні твердої фази. Таким чином, виникає фіксований С-кінець майбутньої поліпептидної молекули. Захист з МН2-групи знімається і додається інша а-амінокислота із захищеною аміногрупою. Утворення пептидного зв'язку відбувається в присутності речовини, що активує карбоксильну групу. Якщо зняти захист з МН2-групи, то синтез пептиду можна здійснювати далі. При цьому відмивання від домішок здійснюють після кожного ступеня на носії. І лише після закінчення синтезу поліпептид знімають з носія. На сьогодні цей синтез здійснюється за допомогою приладів-синтезаторів.
Школою академіка М.М.Шемякіна запропонований так званий рідиннофазний метод синтезу білків, у якому твердий полімерний носій замінений розчинним полістиролом. У цьому методі зберігаються переваги синтезу, проведеного в розчині: більш висока швидкість реакцій і більш широкий вибір реагентів і захисних груп.
5. Водні розчини білків є термодинамічно стійкими і можуть тривалий час перебувати в такому стані, не випадаючи в осад. Іншими словами, вони мають властивості істинних розчинів. Поряд із цим через високу молекулярну масу (понад 4-5 тисяч дальтон) розчинам білка характерні властивості колоїдних розчинів: мала швидкість дифузії, нездатність проходити через напівпроникні мембрани; висока в'язкість, здатність розсіювати промені видимого світла. І все-таки розчини білків не є типовими колоїдними розчинами, які проявляють стабільність за наявності стабілізаторів, що дають змогу запобігати осадженню колоїдів. Коагуляція білків може бути оборотною і необоротною. Подвійність властивостей обумовлена амінокислотним складом білка і структурною організацією.
У випадку розчинення білків у воді, завдяки наявності в них вільних аміно- і карбоксильних груп, розміщених на поверхні молекули білка, обумовлюються кислотно-основні властивості. Кислотні властивості білка залежать від присутності глутамінової та аспарагінової кислот. Менший внесок у кислотність білка роблять тирозин і цистеїн, бо ступінь дисоціації їх незначний. Якщо в молекулі білка переважають кислі амінокислоти, то білки проявляють кислотні властивості і називаються кислими білками (наприклад, фермент шлунка пепсин). При цьому заряд молекул білка позитивний. Лужні властивості білків обумовлені переважанням основних амінокислот (лізин, аргінін, гістидин), через здатність проявляти основні властивості білки називаються основними, а заряд молекул білка негативний.
Заряд молекули білка обумовлює стійкість колоїдних розчинів білка за рахунок відштовхування однойменно заряджених молекул.
Під час розчинення білка відбувається поєднання молекул води з дисоційованими функціональними групами - це не проста адсорбція, а електростатичне поєднання. У результаті утворюється гідратна оболонка. Разом із цим з молекулами води взаємодіють і інші полярні групи бічних радикалів (OH, SH, CONH), з'єднуючись водневими зв'язками, обумовлюючи гідрофільність молекул білка. Крім цього, самі особливості структури білка (вміст у ньому однієї або декількох поліпептидних ланцюгів та їх розміщення в просторі) створюють великий розміг молекули білка, що також сприяє зв'язуванню води. Утворюється гідратна оболонка навколо молекул білка, вона обумовлює стійкість молекул білка в колоїдних розчинах.
У ізоелектричному стані білка відбувається врівноваження негативних і позитивних електричних зарядів - молекула білка електронейтральна. У цьому випадку гідрофільність молекули білка найменша. Якщо гідрофільність молекул з тієї чи іншої причини знижується, то і стійкість молекул зменшується.
Таким чином, до чинників стабільності існування білків у колоїдних розчинах належить наявність електричного заряду білкових частинок і гідратних оболонок (гідрофільність).
6. Як зазначалося вище, розчини білків термодинамічно стійкі і самодовільно не осідають, чинниками стійкості є гідратна оболонка і електричний заряд.
Одним з видів осадження білків з розчинів є процес висолювання, який здійснюється за допомогою концентрованих розчинів солей лужних і нелужних металів. Після додавання такого розчину до розчину білка відбувається дисоціація солей. При цьому, якщо врахувати, що розчини солей концентровані, утворюються у великій кількості іони цих солей, які притягують до себе молекули води з подальшим утворенням гідратної оболонки; за рахунок цього руйнується гідратна оболонка білка, яка забезпечує стійкість білка в розчині. Разом із цим одночасно іони солі адсорбуються на заряджених молекулах білка, знижуючи заряд цих молекул або повністю нейтралізуючи заряд, що сприяє осадженню білка.
Як реагенти для висолювання використовують речовини з різко вираженою полярністю молекул (спирт, ацетон), а також активно гідратуючі іони багатьох сильних електролітів (Na2SO4, (NH4)2SO4 солі магнію і фосфати). Одним з найбільш м'яких дегідратуючих засобів є сульфат амонію. Білки, осаджені цією сіллю, практично не денатуруються. Однак (NH4)2SO4 певним чином зміщує рН розчинів у кислий бік, у результаті чого знижується заряд білкових частинок.
Висолювання білків зазвичай проводять за зниженої температури (до 50С), щоб не викликати денатурацію білків. На розчинність білків під час використання водовіднімаючих розчинників - метилового, етилового спиртів, ацетону - істотно впливають домішки солей, що змінюють рН розчину.
Висолююча дія електролітів залежить від здатності іонів цих електролітів гідратуватися. Ф.Г офмейстером була встановлена така послідовність висолюючої дії аніонів - ліотропний ряд:
![]()
Іони, розташовані ліворуч від Cl-, здатні забирати воду, іони, розташовані праворуч від Cl-, навпаки, підвищують стійкість білків, самі адсорбуються на них, приносячи із собою додаткову воду і заряд. Найбільше висолююча дія характерна для аніону сульфату (SO4-). Катіони розташовуються у ліотропному ряду так:
![]()
На практиці для висолювання білків найчастіше використовують солі амонію і натрію.
Процес висолювання за своєю природою і механізмом істотно відрізняється від коагуляції електролітами. Коагуляція відбувається у випадку невеликих концентрацій електролітів, а висолювання - у дуже концентрованих розчинах. Вплив природи іонів на процеси коагуляції та висолювання протилежний.
Осадження білків проводиться під час висолювання без порушення їх нативної природи. Після відмивання електролітів білки можуть бути знову переведені в розчинений стан.
Існують методи, коли осадження необоротне. Це має місце, коли відбувається глибоке порушення структури молекули білка. Так, у разі підвищення температури руйнується гідратна оболонка молекули білка і змінюється її структура. Унаслідок цього білок втрачає гідрофільність і легко осідає.
Механізм осадження білків під дією солей важких металів (Cu, Pb, Hg, Ag та ін.) відрізняється від механізму процесу висолювання. Іони важких металів взаємодіють з білками, утворюючи нерозчинні у воді комплекси. Тому осадження білка під впливом солей важких металів, також як і в умовах дії високих температур, є процесом необоротним.
7. У нативному стані всі білки мають характерну структуру і вирізняються певними фізико-хімічними та біологічними властивостями. Під впливом зовнішніх чинників - зміна температури, рН-середовища, дія важких металів, опромінення, вплив хімічних речовин - багато білків втрачають свою структуру і властивості, молекула білка переходить у хаотичний стан. У цих умовах білок втрачає нативні специфічні властивості і набуває нових, не характерних для нього. Такі зміни в більшості випадків необоротні або малооборотні і називаються денатурацією білка. Найхарактернішою ознакою денатурації білка є різке зниження або повна втрата білком його біологічної активності. Наприклад, денатурований інсулін втрачає свої специфічні якості гормону, пепсин - здатність перетравлювати білки, а білки-антитіла - свої захисні властивості. При денатурації білків, як правило, не відбувається істотних порушень пептидних зв'язків у поліпептидних ланцюгах, тобто зберігається первинна структура білків. В основі денатурації лежить розрив зв'язків, які стабілізують вищі структури білка (четвертинна, третинна, вторинна). У результаті відбувається розрив поліпептидного ланцюга і білок набуває форми невпорядкованого клубка та випадає в осад. Денатурація характерна тільки для природних білків.
До хімічних реагентів, що викликають денатурацію білка, належать кислоти, луги, алкалоїди, солі важких металів (гідраргірум, купрум, кадмій та ін.). Краще денатуруються білки в дуже сильних кислотах (рН < 2) або лугах (рН > 10-11). Іони важких металів і алкалоїди утворюють з полярними групами білків міцні нерозчинні комплекси, що супроводжується руйнуванням водневих та іонних зв'язків і повною втратою білком нативних властивостей.
Денатурація може бути зворотньою - ренатурацією. Вона призводить до легко відновлюваних змін у структурі і до відновлення біологічних властивостей білка. Для денатурованих білків характерні такі ознаки:
1) порівняно з нативними білками, у них збільшується кількість вільних функціональних груп (-СООН, -NH2, -SH, -OH), тому що частина їх була захована в середині білкової молекули і не виявлялася звичайними методами;
2) зменшення розчинності і випадання в осад (через втрату гідратної оболонки, нейтралізації заряду, підвищення в'язкості, звільнення гідрофобних радикалів);
3) зміна конфігурації білка;
4) втрата біологічної активності.
Виконайте завдання та перевірте правильність їх розв’язання за еталонами відповідей
Завдання № 1
1. Який заряд має молекула нейтрального білка в лужному середовищі?
А. Негативний. В. Позитивний. С. Нейтральний.
2. Які зв'язки практично не руйнуються?
А. Дисульфідні. В. Пептидні. С. Іонні.
3. До складу якого білка входить магній?
А. Каталаза. В. Хлорофіл. С. Пероксидаза.
Еталони відповідей: 1-А; 2-В; 3-В.
Завдання № 2
1. Який з перерахованих білків є глобулярним?
А. Колаген. В. Білок яйця. С. Кератин.
2. Який з перерахованих чинників стабілізує білок у розчині?
A. Величина молекулярної маси.
B. Гідратна оболонка.
C. Концентрація білка в розчині.
3. Необоротне осадження білків у розчині здійснюють під дією:
A. Насичених розчинів солей лужних і лужноземельних металів.
B. Електрофорезу.
C. Високої температури.
Еталони відповідей: 1-В; 2-В; 3-С.
Завдання № 3
1. Який реагент викликає оборотне осадження білків?
A. Хлорид гідраргіруму.
B. Сульфат натрію.
C. Нітрат аргентуму.
2. Який з перерахованих білків належить до складних?
А. Альбумін. В. Гемоглобін. С. Міозин.
3. Основними зв'язками, що стабілізують вторинну структуру білка, є:
А. Дисульфідні. В. Водневі. С. Пептидні.
Еталони відповідей: 1-В; 2-В; 3-В.