Биохимические основы жизнедеятельности организма человека - Волков Н.И., Несен Э.Н. 2000
Биохимические основы жизнедеятельности организма человека
Обмен энергии в организме
Дыхательная цепь
При непосредственном взаимодействии водорода с кислородом одновременно выделяется большое количество энергии, что может привести к гибели клетки. Во избежание прямого взаимодействия водорода с кислородом в клетке во внутренних мембранах митохондрий имеется специальная система переносчиков водорода к кислороду, обеспечивающая постепенное освобождение энергии. Такая система называется электронтранспортной, или дыхательной цепью.
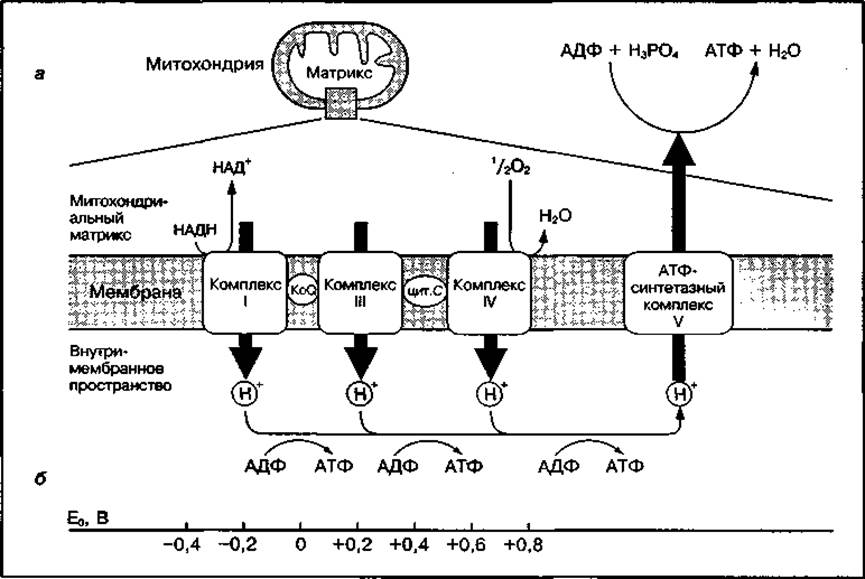
Рис. 19 Последовательность расположения переносчиков электронов в дыхательной цепи и участки, где происходит ресинтез АТФ (а), и изменение окислительно-восстановительного потенциала, вызывающего синтез АТФ (б)
В состав дыхательной цепи входят три основных ферментных комплекса, осуществляющих постепенную передачу электронов от НАДН к атомарному кислороду (рис. 19). Переносчики располагаются в дыхательной цепи в определенной последовательности, взаимосвязанной с их окислительно-восстановительным потенциалом.
Первый комплекс дыхательной цепи, называемый НАД-дегидрогеназным, осуществляет прием электронов от НАДН (в отдельных случаях — от ФАДН2) и передачу их на передатчик убихинон (кофермент Q). В передаче электронов участвуют атомы железа (FeS), которые входят в состав серосодержащих ферментов этого комплекса. Передатчик электронов убихинон представляет собой небольшую жирорастворимую молекулу, подобную витамину К. Он способен принимать один или два электрона и отдавать их. При этом убихинон связывает протоны из водной среды:
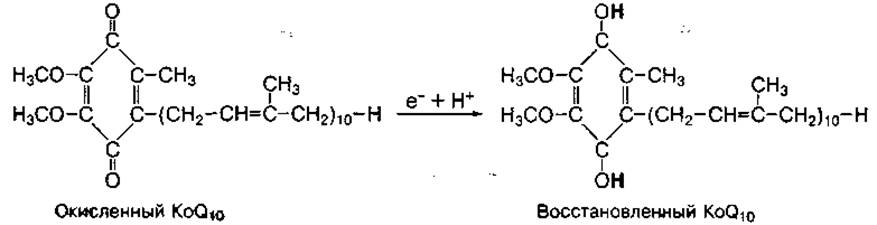
В организме человека находятся преимущественно коферменты Q10, хотя в растениях и животных представлен кофермент Q8. Они не связаны прочно с белками и являются лабильными переносчиками электронов, перемещающимися между компонентами первого и второго комплексов.
Следующий комплекс дыхательной цепи (III) представляет собой ансамбль из двух цитохромов — b и с1 и называется комплексом b—с1 Цитохромы — это группа сложных белков, которые содержат небелковую геминовую группу, близкую по строению к гему белка гемоглобина, в состав которой входит железо, способное отдавать и присоединять электроны:
![]()
Этот комплекс принимает электроны от кофермента Q и передает их на цитохром с — небольшую белковую молекулу, встроенную в мембрану митохондрий, которая переносит эти электроны на дыхательный комплекс IV.
Комплекс IV называется цитохромоксидазным и включает цитохромы а и а3, которые представляют собой сложный фермент, обозначаемый как цитохромоксидаза. Цитохромоксидаза передает электроны с цитохрома с на атомарный кислород. В состав этого комплекса входит медь (Си), которая участвует в передаче электронов. Одновременно из окружающей среды к кислороду присоединяются два протона водорода, в результате чего образуется молекула воды: 1/2O2 + 2е- + 2Н+ → Н2O.
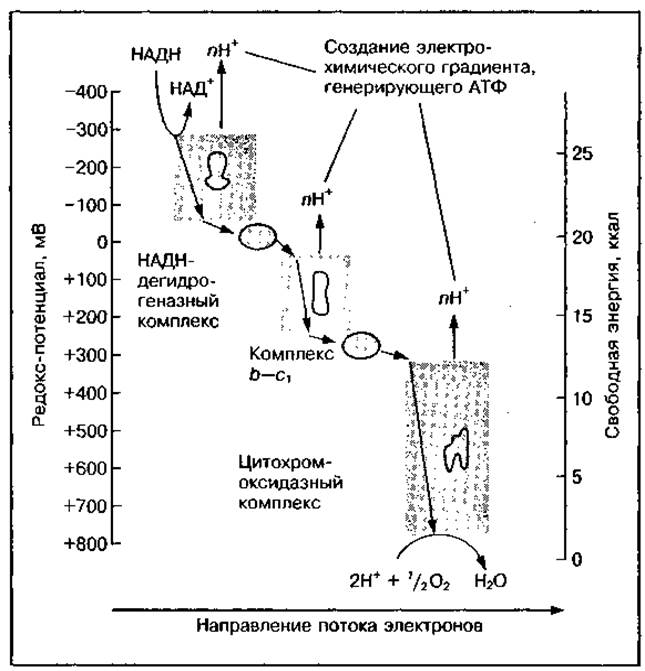
Рис. 20 Снижение окислительновосстановительного потенциала (редокс-потенциала) на различных участках дыхательной цепи при прохождении электрона от НАДН к кислороду
Около 90 % кислорода, поступившего в клетки, используется в этих процессах биологического окисления.
Таким образом, в клетках на внутренней мембране митохондрий находятся дыхательные системы, осуществляющие передачу электронов на кислород, поступающий в организм вместе с вдыхаемым воздухом. Схематически эта цепь может быть представлена в виде
НАДН → НАДН-дегидрогеназный комплекс → убихинон → комплекс b—с1→
→ цитохром с → цитохромный комплекс → 1/22O2
При передаче электронов по дыхательной цепи на кислород происходит постепенное высвобождение энергии. Как видно из рис. 20, окислительно-восстановительный потенциал (редокс-потенциал) в цепи изменяется от -320 мВ (для НАДН) до +820 мВ (для О2).
Резкий перепад потенциала наблюдается между тремя рассмотренными комплексами. Они являются энергопреобразующими устройствами, так ж высвобождаемая свободная энергия используется ими для перекачивания Н+ через внутреннюю мембрану митохондрий в межмембранное пространство (см. рис. 17). Это приводит к образованию электрохимичесго протонного градиента (высокая концентрация Н+ в матриксе — в межмембранном пространстве). Градиент концентрации водорода и мембранный потенциал служат источником энергии для синтеза АТФ в процессе окислительного фосфорилирования и поддержания необходимо уровня АТФ в клетке.