Биохимические основы жизнедеятельности организма человека - Волков Н.И., Несен Э.Н. 2000
Биохимия спорта
Биохимия мыши и мышечного сокращения
Структурные и биохимические изменения в мышцах при сокращении и расслаблении
В расслабленной мышце актиновые нити входят в пространство между миозиновыми нитями по краям дисков А, но не контактируют с ними (см. рис. 115). Центры АТФ-азной активности, находящиеся на головках миозина, присоединяют к себе АТФ, но не расщепляют ее. Для активации АТФ-азы миозина необходимо присутствие ионов Са2+. В саркоплазме покоящейся мышцы концентрация свободных ионов Са2+ очень низкая (10-7 моль ⋅ л-1), так как они находятся в связанном состоянии в пузырьках саркоплазматического ретикулума. АТФ не позволяет контактировать миозиновым нитям с актиновыми. В этом случае АТФ действует как пластифицирующий агент, препятствующий образованию поперечных спаек между актином и миозином. Кроме того, в отсутствие ионов Са2+ молекулы тропонина, расположенные в "овражке" между двумя скрученными полипептидными цепями актина в составе тонких нитей, также блокируют активные центры взаимодействия актина с миозином. Такой двойной ингибирующий эффект препятствует образованию поперечных спаек между толстыми и тонкими нитями в миофибриллах, предохраняет покоящуюся мышцу от бесполезных затрат АТФ и обусловливает упругость (эластичность) в этом состоянии.
Основные биохимические процессы, которые происходят при сокращении мышц, представлены на рис. 119. Сокращение запускается нервным импульсом. При этом в синапсе (1) — месте контакта нервного окончания с сарколеммой выделяется нейропередатчик ацетилхолин (2). Ацетилхолин (Ах) вызывает возбуждение сарколеммы, сопровождающееся деполяризацией мембраны и образованием на ее поверхности потенциала действия (3). Потенциал действия распространяется в глубь волокна через Т-системы, которые контактируют с мембранами саркоплазматического ретикулума. Возбуждение достигает мембранных образований саркоплазматического ретикулума и способствует выходу ионов Са2+ из пузырьков ретикулума в саркоплазму (5). Повышение концентрации свободных ионов Са2+ в области миозиновых нитей активирует АТФ-азные центры в головках миозина. Происходит расщепление АТФ, но продукты этой реакции — АДФ и Фн — остаются на молекуле миозина (6). В таком состоянии миозиновые головки уже способны взаимодействовать с актином, однако центр их взаимодействия блокирован тропонином. В снятии блока и освобождении актиновых центров на поверхности актиновых нитей также участвуют ионы Са2+, которые связываются с тропонином и снимают блок (7). Между головками миозина и активными центрами актина образуются поперечные спайки в виде актомиозинового комплекса. Образование актомиозиновых комплексов стимулирует отщепление АДФ и Фн от головок миозиновых молекул, а выделяющаяся при этом энергия используется для конформационных изменений сократительных белков (8).
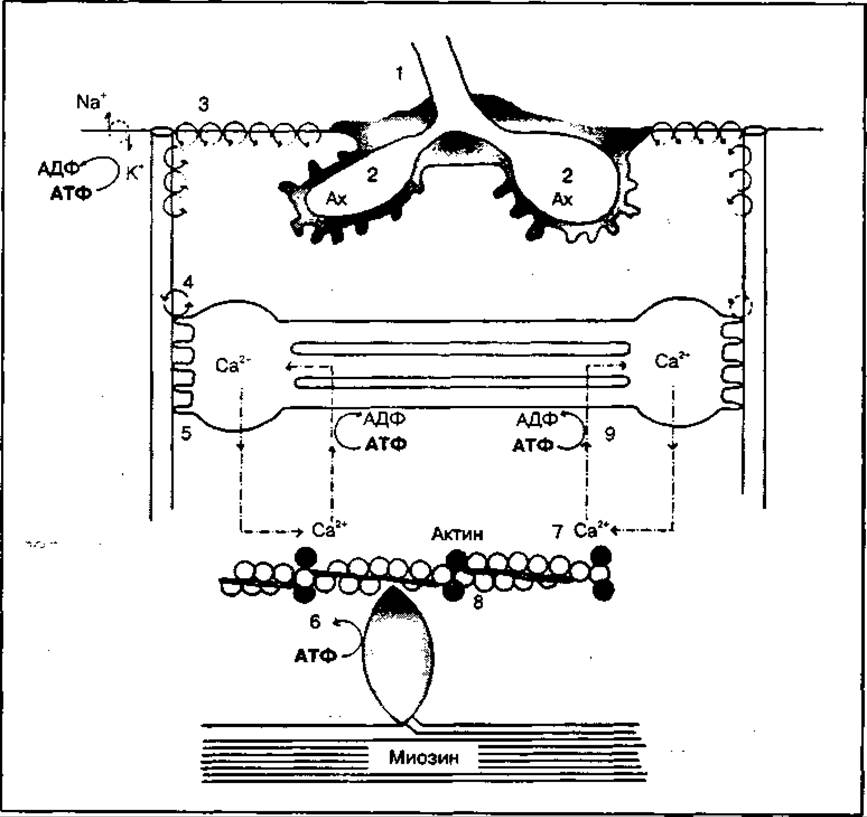
Рис. 119 Основные биохимические процессы, обеспечивающие сокращение мышц
Головки миозиновых молекул изгибаются, принимая стреловидное положение по отношению к оси миозиновой нити, при этом между толстыми и тонкими нитями развивается напряжение, сдвигающее тонкую нить по направлению к центру саркомера. Каждая спайка между актиновыми и миозиновыми нитями в процессе сокращения действует независимо от образования других спаек.
Общий процесс сокращения, проявляющийся в укорочении мышечного волокна и развитии напряжения, является результатом суммирования одновременного образования большого числа спаек по всей длине миофибрилл, вовлеченных в процесс сокращения возбужденной мышцы. Величина напряжения в сокращающейся мышце будет пропорциональна количеству поперечных спаек или площади их наложения в пределах каждого саркомера, что диктуется нервной системой (рис. 120). При значительном растяжении мышцы (длине саркомера более 3,65 мкм) тонкие нити полностью выходят за пределы дисков А и напряжение в мышце отсутствует. По мере вхождения тонких нитей между толстыми и увеличения площади их наложения друг на друга напряжение в мышце постепенно увеличивается, достигая максимума при длине саркомера от 2,25 до 2,00 мм. При более значительном сокращении мышцы тонкие нити перекрываются в центре дисков А и сжимаются в зоне Н, образуя полосу сокращения. Напряжение мышцы в этой стадии сокращения быстро снижается.
Рассмотренные процессы мышечного сокращения согласуются с теорией "скольжения" филаментов, выдвинутой английским биофизиком X. Хаксли (1968).
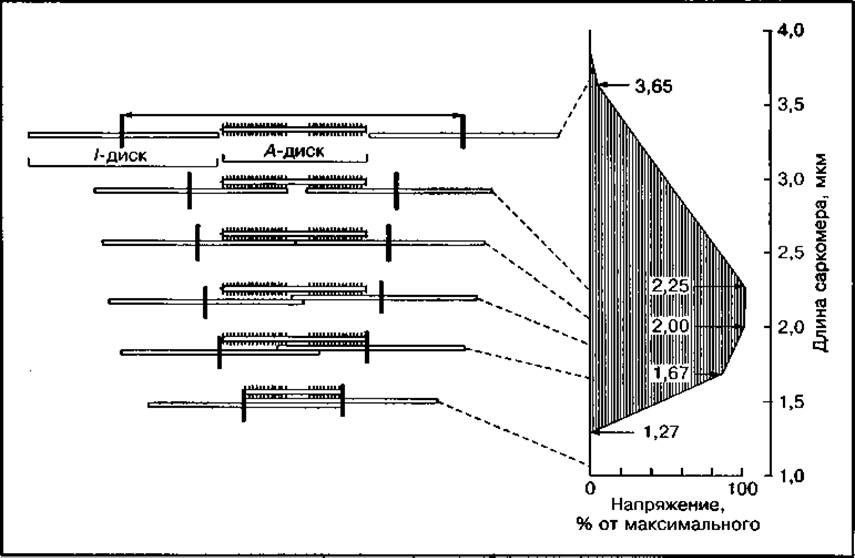
Рис. 120 Изменение длины саркомера и развития напряжения в результате суммирования образовавшихся спаек в процессе сокращения