Основы биохимии - А. А. Анисимов 1986
Обмен белков и аминокислот
Фиксация молекулярного азота воздуха
Подсчитано, что на Земле содержится около 3,8х1015 т азота (в расчете на элементарный). Основную часть его составляет молекулярный азот атмосферы — N2. Однако человек, животные и высшие растения не способны самостоятельно усваивать такой азот. Тем не менее все живые организмы для нормального функционирования должны потреблять азот в большом количестве; азотсодержащие соединения — белки и аминокислоты — являются важнейшими компонентами пищевого рациона человека и животных, столь же важны и необходимы минеральные соли азота для питания высших растений.
Обеспечение населения пищевым белком — одна из наиболее острых проблем, стоящих в настоящее время перед человечеством, особенно в развивающихся, бывших колониальных странах. Этой проблеме в наши дни уделяется столь большое внимание, что при Организации Объединенных Наций создан специальный комитет по всестороннему изучению проблемы белковых ресурсов и разработке практических рекомендаций по предотвращению белкового голода. Д. И. Менделеев, оценивая эту проблему, писал, что вопрос о способах превращения азота воздуха в почвенные азотистые соединения или в ассимилируемый азот, способный поглощаться растениями и давать в них сложные (белковые) вещества, составляет один из таких вопросов, которые представляют великий теоретический и практический интерес. По мнению акад. С. П. Костычева, азотное голодание является тем фактором, который преимущественно перед всеми остальными ограничивает развитие жизни на Земле и задерживает размножение организмов.
При производстве азотных минеральных удобрений азот воздуха превращают промышленным путем в аммиак или азотную кислоту. Однако этот азот составляет только 2—3% от азота, содержащегося в урожае сельскохозяйственных растений всей Земли. Практически почти весь азот, находящийся в живых организмах нашей планеты, имеет своим источником азот атмосферы, фиксированный микроорганизмами, ибо только они способны к самостоятельному усвоению молекулярного азота.
Некоторые высшие растения фиксируют атмосферный азот, вступая в симбиотические отношения с бактериями своих корневых клубеньков, и таким путем вносят значительный вклад в общее усвоение молекулярного азота. Считают, что из известных 13 000 видов бобовых растений большинство способны симбиотически фиксировать азот, причем в значительных количествах, особенно это относится к культурным бобовым растениям (горох, соя и др.). Примерно 250 видов других семейств, небобовых, тоже способны к симбиотической фиксации азота (например, ольха, лисохвост, облепиха).
Бактерии клубеньковых бобовых растений принадлежат в основном к роду Rhizobium. Из свободноживущих бактерий фиксируют молекулярный азот почвенные бактерии рода Azotobacier (аэроб), Clostridium (анаэроб) и некоторые факультативные анаэробы, а также все фотосинтезирующие бактерии. На долю сине- зеленых водорослей приходится 10—15% всего количества фиксируемого N2атмосферы.
При промышленном получении аммиака из N2 затрачивается большое количество энергии, даже в присутствии катализаторов требуется повышение температуры до +500°С и давления до 300— 350 атм. Поэтому не может не восхищать способность микроорганизмов осуществлять практически тот же процесс при обычном атмосферном давлении и невысоких температурах. Основной «секрет» этой способности — наличие у микроорганизмов особой ферментативной системы фиксации азота. Поэтому не удивительно, что во многих лабораториях мира в настоящее время очень активно исследуют фиксацию N2, и крайне заманчивой, интересной и важной является попытка решить вопрос об имитации в промышленных условиях ферментативных процессов, протекающих у азотфиксирующих микроорганизмов. Большие и интересные работы по биохимии усвоения молекулярного азота воздуха проведены в Советском Союзе под руководством В. Л. Кретовича в Институте биохимии АН СССР.
Все самые разнообразные азотфиксирующие микроорганизмы содержат одинаковую ферментативную систему, катализирующую превращение молекулярного азота в аммиак. Эта система получила название нитрогеназы. Кроме того, для фиксации N2 необходимы сильные восстановители (поток электронов), АТФ и Mg2+. Природа доноров электронов различна у разных микроорганизмов. У аэробных бактерий (Azotobacier, Rhizobium) необходимые для фиксации N2 восстановители и АТФ образуются в ходе углеводного обмена, в реакциях с участием НАДФ. Фотосинтетические бактерии и сине-зеленые водоросли способны к фотохимическому образованию сильных восстановителей.
Нитрогеназа (КФ 1.18.2.1) состоит из двух белков—Mo-Fe-белка (молибдоферредоксин) и Fe-белка (азоферредоксин). Молекулярная масса первого (в зависимости от вида микроорганизмов) составляет от 200 000 до 250 000; это тетрамер, содержащий два атома молибдена, негемовое железо и лабильный сульфид. Молекулярная масса Fe-белка составляет от 50 000 до 70 000; он является димером, тоже содержит негемовое железо и лабильный сульфид (рис. 5.1).
В процессе фиксации АТФ взаимодействует с азоферредоксцном. При этом выделяется АДФ, а азоферредоксин претерпевает конформационную перестройку, вследствие чего его окислительно-восстановительный потенциал понижается с —280 до —400 мВ. Азоферредоксин становится сильным восстановителем, передает электроны на молибдоферредоксин, где и осуществляется восстановление N2до NH3. Суммарно процесс фиксации азота можно выразить реакцией
N2+ 6е- + 12АТФ + 12Н2О → 2NH+4+ 12АДФ + 12Н3РО4 + 4Н+
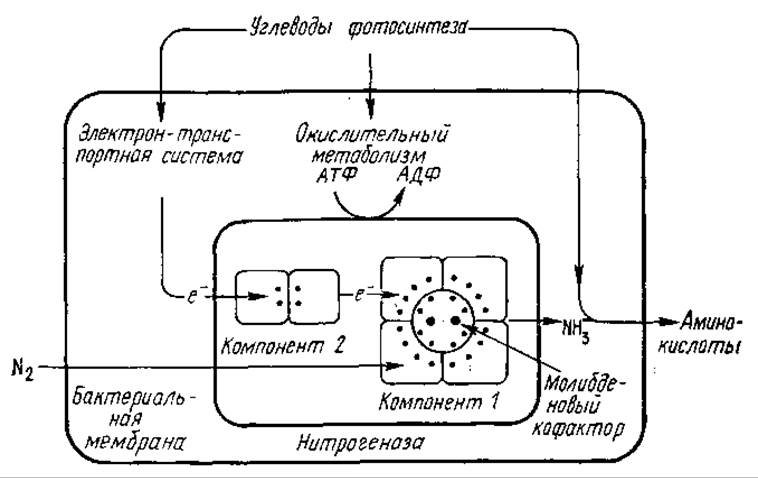
Рис. 5.1. Схема биологической фиксации азота
Нитрогеназа обладает широкой субстратной специфичностью, кроме N2она может восстанавливать цианиды, закись азота, ацетилен и др. Нитрогеназа при участии АТФ катализирует также восстановление ионов водорода с образованием молекулярного водорода. У некоторых азотфиксаторов этот процесс протекает одновременно с азотфиксацией. Предполагают, что данное свойство азотфиксирующих микроорганизмов в будущем можно будет использовать для получения дешевого топлива в виде молекулярного водорода.
Для азотфиксации очень важен молибден, функции его в этом процессе разнообразны. Он поддерживает определенную конформацию молекулы нитрогеназы, участвует в связывании азота и переносе электронов, индуцирует синтез нитрогеназы. Молекула газообразного азота очень прочна, для разрыва трех связей, соединяющих ее атомы, требуется около 940 кДж/моль.
Существенным вопросом в процессе азотфиксации является потребление достаточно большого количества энергии, необходимой для разрыва прочных внутримолекулярных связей N2при его восстановлении до аммиака. Azotobacter, например, для связывания 15 мг азота расходует на дыхание 1 г сахара, а Clostridium pasteurianum еще больше (5—6 г сахара). У клубеньковых бактерий энергетическим материалом являются продукты фотосинтеза, поступающие в корневую систему из листьев. Часть этих ассимилятов может перерабатываться и откладываться в запас в бактероидах в форме поли-β-гидроксимасляной кислоты (ПОМ).
Способность бактероидов к азотфиксации связана и с особенностями их окислительно-восстановительных систем, в частности с присутствием особого гемопротеина — легоглобина. Предполагают, что он, подобно гемоглобину животных, выполняет роль переносчика кислорода к бактероидам, находящимся в клубеньке в условиях затрудненного доступа кислорода. Образовавшийся в результате азотфиксации аммиак в дальнейшем ассимилируется в основном в растительной ткани клубенька. Первый этап этой ассимиляции заключается в связывании аммиака кетокислотами с образованием аминокислот.