Основы биохимии - А. А. Анисимов 1986
Углеводы
Образование углеводов в процессах фотосинтеза и хемосинтеза
6.5.1. Фотосинтез. Фотосинтез — это совокупность процессов, в ходе которых запасается солнечная энергия в виде химических связей органических соединений, синтезируемых из неорганических веществ.
1 Этот раздел излагается очень сокращенно, так как в соответствии с программой Министерства высшего и среднего специального образования РСФСР фотосинтез должен подробно рассматриваться в лекционных курсах и учебниках по физиологии растений.
Фотосинтез состоит из двух фаз: световой (фотофизический и фотохимический этапы) и темновой. В ходе световой фазы происходит поглощение солнечной энергии хлорофиллом и передача ее в «реакционный центр», где в результате химических реакций, включающих транспорт электронов между различными переносчиками и сопряженного с ним фосфорилирования, образуются восстановительные и энергетические эквиваленты (НАДФН и АТФ). Для нормального функционирования реакций световой фазы кроме света необходимы хлорофилл и вода (или другой источник водорода). Различные хлорофиллы способны поглощать кванты света с определенной длиной волны и переходить при этом в возбужденное состояние. Переход в исходное состояние приводит к высвобождению энергии, которая через ряд промежуточных стадий запасается в форме АТФ и НАДФН.
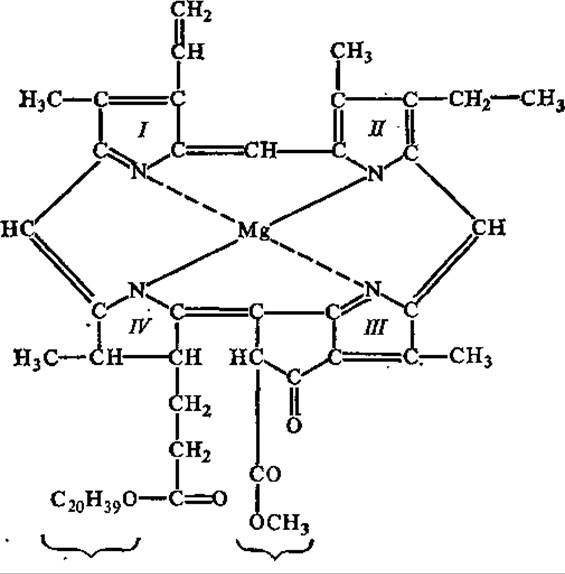
Рис. 6.6. Строение хлорофилла а:
S—IV — пиррольные циклы; пунктиром показаны координационные связи
Все известные в настоящее время фотосинтезирующие организмы содержат хлорофиллы — зеленые магний-порфириновые пигменты. Известно свыше десяти их видов, различающихся природой химических групп, присоединенных к пиррольным структурам порфиринового ядра, окраской, распространением среди живых организмов. Так, у всех зеленых растений содержатся хлорофиллы а и b (рис. 6.6), в диатомовых водорослях — хлорофилл с, в красных водорослях — хлорофилл d. В клетках пурпурных бактерий обнаружены бактериохлорофиллы а и b, а в зеленых бактериях — бактериохлорофиллы с и d. Важным свойством молекул хлорофилла является их способность взаимодействовать с белками и друг с другом, образуя агрегированные формы с различными спектрами поглощения.
Наряду с зелеными пигментами в хлоропластах и хроматофорах содержатся каротиноиды — желтые и оранжевые пигменты полиизопреновой природы. Каротиноиды могут быть разделены на ряд групп по своему строению: собственно каротиноиды, гидроксилсодержащие каротиноиды и каротиноиды, содержащие карбонильные группы. Основными представителями у высших растений являются ß-каротин и ксантофилл (рис. 6.7). Предполагают, что каротиноиды, используя лучи, не поглощаемые хлорофиллом, передают их энергию на молекулы хлорофилла. Существуют данные, что каротиноиды предохраняют молекулы хлорофилла от разрушения в процессе фотоокисления, а также играют определенную роль в половом процессе при прорастании пыльцы и росте пыльцевых трубок, у высших растений и при созревании половой клетки у водорослей и грибов.
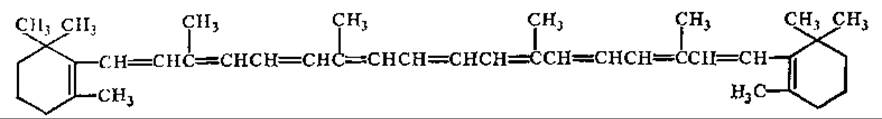
Рис. 6.7. Структура ß-каротина
Третья группа пигментов — фикобилины. Это красные и синие пигменты (фикоэритрины, фикоцианины), содержащиеся в хроматофорах некоторых водорослей. В основе химического строения фикобилинов лежит та же тетрапиррольная структура, но пиррольные группы расположены линейно. Фикобилины поглощают энергию света в зеленой и желтой областях спектра и передают ее на молекулу хлорофилла, после чего она используется в процессе фотосинтеза. Наличие фикобилинов у водорослей — пример приспособления в ходе эволюции к поглощению тех лучей солнечного света, которые проникают через толщу морской воды. Оптические свойства хлорофилла, его роль как фотосенсибилизатора1 были впервые исследованы великим русским физиологом К. А. Тимирязевым. В дальнейшем крупный вклад в решение этой проблемы был сделан А. Н. Терениным и А. А. Красновским.
1 Фотосенсибилизаторы — вещества, поглощающие энергию света и передающие ее на ту или иную бесцветную молекулу.
Темповая фаза фотосинтеза — это фиксация и восстановление СО2 с образованием углеводов и других конечных продуктов фотосинтеза. На этой стадии свет не нужен, а используются образованные в световой фазе восстановительные и энергетические эквиваленты. Во-время темновой фазы атомы водорода, поставляемые световыми реакциями, используются для восстановления СО2 до углеводов согласно общему уравнению фотосинтеза:
![]()
При этом на каждый моль синтезированного углевода запасается ~160 кДж энергии.
Процесс восстановления СО2 начинается с его присоединения к пятиуглеродному акцептору рибулозо-1,5-бисфосфату (РуБФ). Образующееся шестиуглеродное соединение очень нестойко. Опытами М. Кальвина с меченым оксидом углерода (IV) установлено, что при длительности фотосинтеза 2 с первым фиксируемым меченым соединением является не С6-соединение, а 3-фосфоглицериновая кислота (ФГК), причем метка обнаруживается только в карбоксиле. Реакция карбоксилирования катализируется рибулозобисфосфат-карбоксилазой (РБФ-карбоксилаза). Затем образующаяся ФГК фосфорилируется при участии фермента фосфоглицераткиназы с использованием АТФ и превращается в 1,3-бисфосфо-D-глицериновую кислоту, которая более реакционноспособна и легче восстанавливается в глицеральдегид-3-фосфат (ГАФ).
В реакции восстановления принимают участие образовавшийся в световой стадии НАДФН и фермент глицеральдегидфосфатдегидрогеназа. Часть образовавшихся молекул ГАФ под действием фермента триозофосфатизомеразы превращается в дигидроксиацетонфосфат (ДГАФ). Эти два триозофосфата конденсируются под действием альдолазы. Сначала образуется фруктозо-1,6-дифосфат (ФДФ), далее — монофосфаты фруктозы (Ф6Ф) и глюкозы (Г6Ф) и, наконец, — сахароза и крахмал. Для того чтобы процесс фотосинтеза продолжался, необходима постоянная регенерация РуБФ, поэтому в одном обороте цикла участвуют 6 молекул РуБФ, которые фиксируют 6 молекул СO2. Образующиеся 12 молекул ФГК, а после ее восстановления 12 молекул ГАФ расходуются следующим образом:
5ГАФ → 5ДГАФ
ЗГАФ + 3ДГАФ → 3ФДФ → 3Ф6Ф
2ГАФ + 2Ф6Ф → 2 Ксилулоза-5Ф + 2 Эритрозо-4Ф
2ГАФ + 2 Седогептулозо-7Ф → 2 Ксилулозо-5Ф + 2 Рибозо-5Ф
Таким образом, две молекулы глицеральдегид-3-фосфата образуют одну молекулу гексозы, которая выходит из цикла, а 10 идут на регенерацию шести молекул рибулозо-1,5 бисфосфата, и цикл замыкается.
На рис. 6.8 представлены все реакции темновой фазы фотосинтеза. Большая заслуга в разработке этой схемы принадлежит американскому биохимику М. Кальвину и его сотрудникам (1957), поэтому она носит название цикла Кальвина.
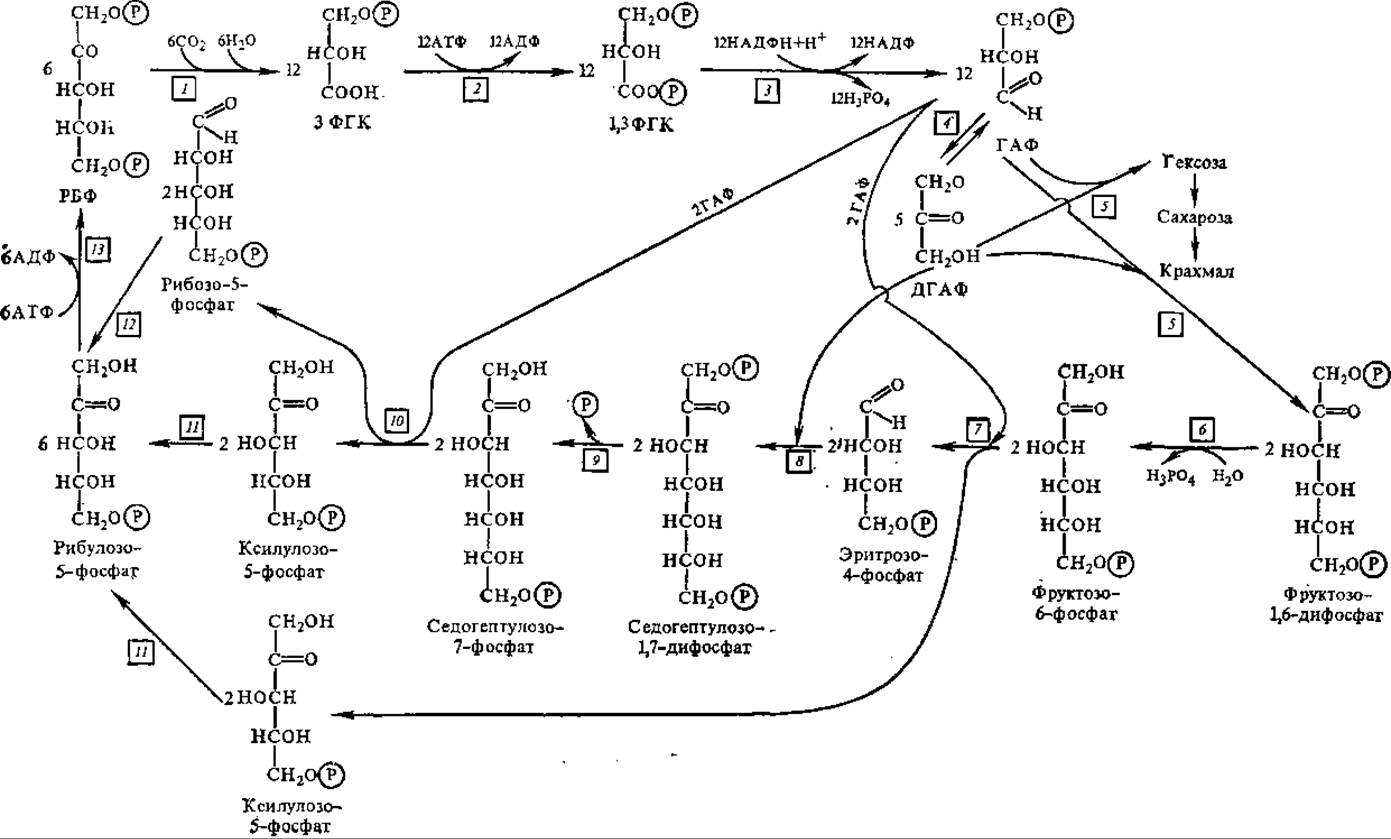
Рис. 6.8. Биохимические превращения углерода при фотосинтезе (цикл Кальвина):
цифрами указаны ферменты, катализирующие эти превращения; 1 — рибулозобисфосфат-карбоксилаза, 2 — фосфоглицераткиназа, 3 — глицеральдегидфосфатдегидрогеназа, 4 — трнозофосфатазомераза, 5 — альдолаза, 6 — фосфатаза, 7 — транс-кетолаза, 3 — альдолаза, 9 — фосфатаза, 10 — транскетолаза, 11 — фосфокетопентозоэпимераза, 12 — рибозофосфат-изомераза, 13 — фосфорибулокиназа; остальные обозначения см. в тексте.
На схеме видны три пути, ведущих к регенерации РуБФ, что обеспечивает непрерывный приток этого соединения, поэтому затруднений с фиксацией СО2 не возникает. Суммарное уравнение цикла Кальвина имеет следующий вид:
6РуБФ + 6СО2 + 12НАДФН + 18АТФ + 12Н+ → 6РуБФ +
+ 12НАДФ+ + 18АДФ + 18Н3РО4+ Гексоза
Долгое время считали, что единственный органический продукт фотосинтеза — углеводы, а все другие метаболиты образуются в растениях из углеводов в результате реакций, не связанных с фотосинтезом. Однако еще в начале XX в. известные русские физиологи растений В. В. Сапожников и Ф. Н. Крашенинников указывали на возможность образования в процессе фотосинтеза аминокислот и белков, приводя в доказательство убедительные аргументы.
Экспериментальные доказательства образования аминокислот при фотосинтезе (с использованием меченых атомов) были получены в СССР А. А. Ничипоровичем и сотрудниками. В настоящее время не вызывает сомнений представление о том, что в хлоропластах в качестве непосредственных продуктов фотосинтеза кроме углеводов синтезируется целый ряд других соединений. Уже самый первый продукт фотосинтеза — ФГК — путем аминирования может превращаться в серин. ФГК легко преобразуется в фосфоенолпировиноградную кислоту (ФЕП), затем в пируват, который может служить исходным продуктом для образования ряда органических кислот; его аминирование дает аланин.
Карбоксилйрование ФЕП приводит к синтезу оксалоацетата с возможным восстановлением его до яблочной или аминированием до аспарагиновой кислот. При взаимодействии ФЕП с другим промежуточным продуктом цикла Кальвина — эритрозофосфатом образуется циклическая шикимовая кислота, являющаяся исходным соединением для биосинтеза ароматических аминокислот, а также ряда фенолов, дубильных веществ, гликозидов и др.
У некоторых растений в качестве первичных продуктов фотосинтеза сначала образуются оксалоацетат и малат в результате карбоксилирования ФЕП под действием ФЕП-карбоксилазы. По-скольку первичные продукты в этом случае содержат четыре атома углерода, его называют С4-путь фотосинтеза в отличие от цикла Кальвина, где образующаяся ФГК содержит три атома углерода (С3-путь). С4-Путь фотосинтеза, в котором принимают участие два типа клеток и два типа хлоропластов, называется кооперативным. Он был открыт и исследован в 60-х годах XX в. Ю. С. Карпиловым (СССР), М. Хэтчем и К. Слэком (Австралия).
6.5.2. Хемосинтез. Гетеротрофная фиксация СО2. Известный русский микробиолог С. Н. Виноградский показал (1885—1895), что органические вещества синтезируются в природе не только путем фотосинтеза в зеленых растениях, но и бактериями, не содержащими хлорофилла. Энергию, необходимую для синтеза органических соединений, эти бактерии получают при окислении различных неорганических соединений: Fe, N, S, Н, Sb, Mn. Этот процесс называется хемосинтезом. Некоторые хемосинтетики используют в качестве доноров водорода простейшие органические вещества — метан, метанол и пр.
Исследование химизма ассимиляции меченого оксида углерода (IV) (14СО2) различными хемосинтезирующими бактериями показало, что первым стойким продуктом хемосинтеза является фосфоглицериновая кислота, а присоединение СО2 к рибулозобисфосфату, т. е, цикл Кальвина, — основным механизмом ассимиляции СО2. У многих хемосинтезирующих бактерий цикл Кальвина — главный, но не единственный путь образования органических веществ.
Таким образом, процессы фотосинтеза и хемосинтеза — источники органического вещества на Земле.
А. Ф. Лебедевым еще в 1914 г. была высказана мысль, что гетеротрофные организмы могут частично ассимилировать углерод не только из готовых органических соединений, но и из СО2, связывая последний с некоторыми кетокислотами. В последние годы это получило экспериментальное подтверждение в исследованиях А. Л. Курсанова, Г. Вуда, С. Очоа и др.
Однако, как уже указывалось, между автотрофами и гетеротрофами существует коренное отличие: первые способны синтезировать органическое вещество полностью за счет неорганических веществ (СО2 и Н2О), вторые усваивают СО2, используя готовое органическое соединение, например пируват.