Биологическая химия - Березов Т. Т., Коровкин Б. Ф. 1998
Биосинтез белка
Трансляция и общие требования к синтезу белка в бесклеточной системе
Рибосомы
Как известно, живые организмы в зависимости от структуры клеток делятся на две группы — прокариоты и эукариоты. Первые не содержат ограниченного мембраной ядра и митохондрий или хлоропластов; они представлены главным образом микроорганизмами. Клетки эукариот животных и растений, включая грибы, напротив, содержат ядра с мембранами, а также митохондрии (в ряде случаев и хлоропласты) и другие субклеточные органеллы.
Оба типа клеток имеют рибосомы, причем рибосомы эукариот (мол. масса 4,2 ∙ 106) значительно большего размера (23 нм в диаметре), чем рибосомы прокариот (мол. масса 2,5 ∙ 106, 8 нм в диаметре). Обычно рибосомы характеризуют по скорости их седиментации в центрифужном поле, которая количественно выражается константой седиментации s в единицах Сведберга S (см. главу 1). Величина s зависит не только от размера частиц, но и от формы и плотности, так что она непропорциональна размеру. Число рибосом в микробной клетке равно примерно 104, а эукариот — около 105.
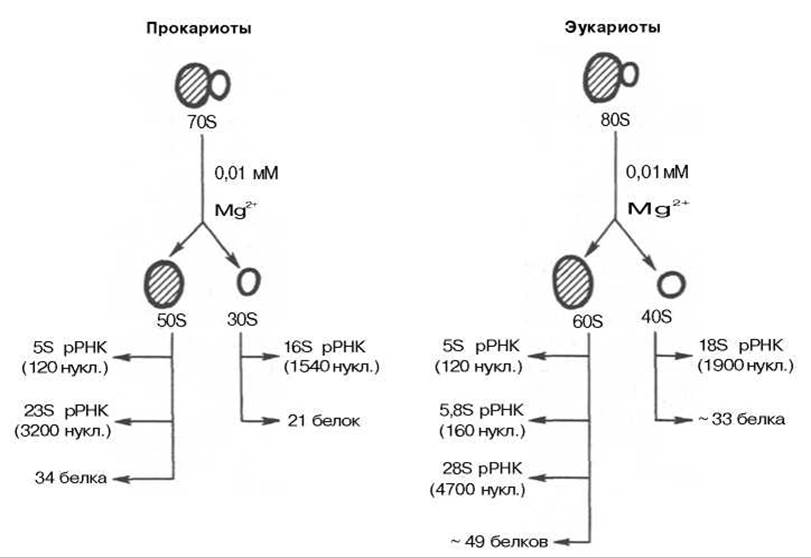
Рис. 14.2. Компоненты рибосом прокариот и эукариот (схема).
Химически рибосомы представляют собой нуклеопротеины, состоящие из РНК и белков, причем 80S рибосомы эукариот содержат примерно равное их количество, а у 70S рибосом прокариот соотношение РНК и белка составляет 65% и 35% соответственно (рис. 14.2). РНК рибосом принято называть рибосомными и обозначать рРНК. Как 80S, так и 70S рибосомы состоят из двух субчастиц, которые можно увидеть под электронным микроскопом или после обработки рибосом растворами, содержащими низкие концентрации ионов Mg2+. При этих условиях рибосомы диссоциируют на субчастицы; последние могут быть отделены друг от друга методом ультрацентрифугирования. Одна из субчастиц по размерам в 2 раза превышает вторую. Так, у 70S рибосом величины s для субчастиц равны 50S и 30S, у 80S рибосом — соответственно 60S и 40S (см. рис. 14.2). Укажем также, что у Е. coli большая и малая субчастицы содержат 34 белка и 21 белок соответственно и, кроме того, 2 молекулы рРНК с коэффициентами седиментации 23S и 5S в большой и одну молекулу рРНК (16S) в малой субчастице. Рибосомные белки не только все выделены, но и секвенированы; отличаются большим разнообразием молекулярной массы (от 6000 до 75000). Считается, что все 55 бактериальных рибосомных белков участвуют в синтезе полипептидов в качестве ферментов или структурных компонентов, но, за исключением небольшого числа, детальная функция большинства из них не выяснена. РНК 23S и 5S содержат 3200 и 120 нуклеотидов соответственно, a 16S РНК — 1540 нуклеотидов. Субчастицы рибосом клеток эукариот построены более сложно. В их составе четыре разные рРНК и более 70 разных белков в обеих субчастицах, при этом большая субчастица (60S) содержит три разного размера рРНК: 28S (4700 нуклеотидов), 5,8S (160 нуклеотидов) и 5S (120 нуклеотидов) — и около 49 белков. Малая субчастица (40S) содержит всего одну молекулу 18S рРНК и около 33 белков. Укажем также, что биологические функции компонентов эукариотических рибосом также связаны, вероятнее всего, с синтезом полипептидной цепи, но их конкретная роль недостаточно раскрыта.
Рибосомы представляют собой сложную молекулярную «машину» («фабрику») синтеза белка. Для выяснения тонких механизмов синтеза белка в рибосомах необходимы более точные сведения о структуре и функциях всех компонентов рибосом. В последнее время получены данные, свидетельствующие о вероятной пространственной трехмерной структуре как целых рибосом, так и их субчастиц. В частности, выяснено, что форму и размеры 30S и 40S субчастиц рибосом предопределяют не белковые молекулы этих частиц, а третичная структура входящих в их состав 16S и 18S рРНК. Более того, по данным акад. А.С. Спирина, для сохранения пространственной морфологической модели всей 30S субчастицы оказалось достаточным наличие только двух белков (из 21), содержащихся в определенных топографических участках молекулы 16S рРНК.
Известно, что рРНК образуется из общего предшественника всех типов клеточных РНК, в свою очередь синтезирующегося на матрице ДНК в ядре (см. главу 13). Рибосомные белки имеют цитоплазматическое происхождение, затем они транспортируются в ядрышки, где и происходит спонтанное образование рибосомных субчастиц путем объединения белков с соответствующими рРНК. Объединенные субчастицы вместе или врозь транспортируются через поры ядерной мембраны обратно в цитоплазму, где группа рибосом вместе с мРНК образует полисомы или полирибосомы, принимающие непосредственное участие в синтезе белка.