Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ I. БІОМОЛЕКУЛИ ТА КЛІТИННІ СТРУКТУРИ
ГЛАВА 4. ВУГЛЕВОДИ ТА ЇХ ПОХІДНІ
4.4. ПЕПТИДОІЛІКАНИ КЛІТИННОЇ СТІНКИ МІКРООРГАНІЗМІВ
Більшість мікробів значну частину свого життєвого циклу проводять у вигляді одноклітинних організмів, тому їх крихка цитоплазматична мембрана обгорнена зовнішньою клітинною стінкою, що захищає мембрану від пошкоджуючих факторів середовища.
Основу клітинної стінки бактерій складають особливі гетерополісахариди, що сполучені з мембранними білками у вигляді протеогліканів або пептидогліканів.
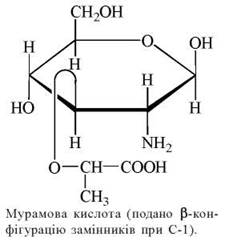
Мурамін — гетерополісахарид клітинної стінки бактерій, нерозгалужені ланцюги якого складаються із залишків N-ацетилглюкозаміну та N-ацетилмурамової кислоти, що сполучені між собою β-1,4-глікозидним зв’язком. Мурамова кислота є похідним D-глюкозаміну, до якого в 3-му положенні приєднана молочна кислота (лактильний залишок).
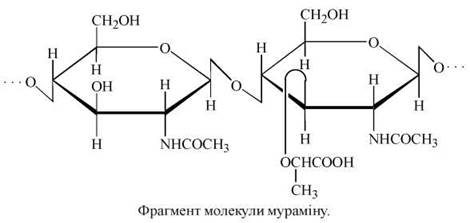
Організація будови мураміну подібна до такої целюлози і хітину (нерозгалужені ланцюги з β-зв’язками між мономерами), що визначає механічну міцність цих гетерополісахаридів та їх структурну функціональну роль. До того ж шлунково-кишковий тракт організму людини не містить ферментів, що здатні розщеплювати ці полісахариди.
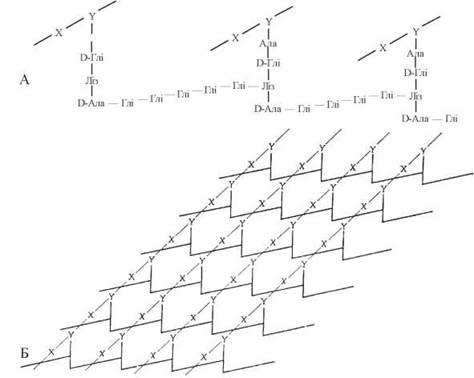
Рис. 4.2. Схема молекулярної організації пептидоглікану бактеріальної стінки — на прикладі муреїну Salmonella aureus (за А. Ленінджером, 1974). А. Будова поперечних пептидних зв’язків між окремими ланцюгами полісахаридів (X — залишок N-ацетилглюкозаміну; Y — залишок N-ацетилмурамової кислоти). Тетрапептидні (L-Ала - D-Глн - L-Ліз - D-Ала) бічні ланцюги кожного полісахаридного (мурамінового) ланцюга сполучені між собою пентапептидними «місточками» з п’яти залишків гліцину. Б. Паралельні гетерополісахаридні ланцюги муреїну зв’язані між собою в єдиний макромолекулярний каркас пептидоглікану за рахунок поперечних пептидних ланцюгів.
Муреїн. У формуванні клітинної стінки бактерій бере участь мурамін, сполучений із пептидними ланцюгами у вигляді протеоглікану (або пептидоглікану) муреїну. Муреїн — це мішаний біополімер, що складається з паралельних полісахаридних ланцюгів, які зв’язані один з одним у поперечному напрямку короткими пептидними ланцюгами. Фактично мурамін можна собі уявити як єдину молекулу, що у вигляді гігантського мішка або каркаса охоплює бактеріальну клітину.
Грампозитивні та грамнегативні мікроорганізми
Стінки бактеріальних клітин, окрім охарактеризованої вище пептидогліканової сітки, мають у своєму складі деякі додаткові біохімічні компоненти полісахаридної, ліпідної та білкової природи, що варіюють у різних мікроорганізмів.
Особливістю складу клітинних стінок є молекулярна основа класичного розподілу бактерій на грампозитивні та грамнегативні, що відрізняються своїм відношенням до забарвлення сумішшю генціанового фіолетового, йоду та сафраніну. Грампозитивні бактерії за умов їх послідовної обробки зазначеними реактивами дають синьо-фіолетове забарвлення внаслідок утримання комплексу йоду з генціановим фіолетовим, грамнегативні бактерії — не дають позитивної реакції на зазначений барвник.
Грампозитивні мікроорганізми — бактерії, характерною особливістю біохімічного складу клітинної стінки яких є наявність тейхоєвих кислот, вплетених у пептидоглікановий матрикс. Тейхоєві кислоти є складними полімерами, ланцюги яких побудовані із залишків спиртів гліцерину та рибіту, що сполучені фосфоді-фірними зв’язками. У деяких тейхоєвих кислотах з основним ланцюгом молекули зв’язані залишки D-аланіну та D-глюкози або N-ацетил-D-глюкозаміну. Пептидоглікани складають більшу частину (40-90 %) маси клітинної стінки грампозитивних бактерій.
Грамнегативні мікроорганізми — бактерії, до складу клітинної стінки яких входить додаткова зовнішня мембрана. Зовнішня мембрана складається із шару ліпополісахаридів, ліпідів та білків, під якими міститься шар пептидогліканів, які, в свою чергу, вкривають внутрішню (цитоплазматичну) мембрану. Загальний вміст пептидогліканів у стінці грамнегативних бактерій значно нижчий (5-10 %), ніж у грампозитивних організмів.
Антибіотики — інгібітори синтезу клітинної стінки
Розщеплення або порушення синтезу окремих компонентів пептидогліканової структури клітинних стінок мікроорганізмів є молекулярною основою антибактеріальної дії деяких антибіотиків.
Пеніциліни — антибіотики, що продукуються пліснявими грибами штамів Penicillium і мають значну бактерицидну активність відносно грампозитивних мікроорганізмів. Механізм біохімічної дії пеніцилінів полягає в гальмуванні однієї із заключних стадій у формуванні пептидоглікану, а саме — взаємодії пентагліцину з термінальним D-аланіном бічних тетрапептидних ланцюгів, що зв’язані з гетеро- полісахаридом. Така дія антибіотика призводить до порушення формування муреїнового каркаса клітинної стінки мікроорганізму, що в остаточному підсумку спричиняє загибель бактерій.
Подібно до пеніцилінів діють цефалоспорини — антибіотики, що виділені з гриба Cephalosporinum acremonium. Цефалоспорини також спричиняють пригнічення активності ферментів, які необхідні для формування пептидогліканів клітинної стінки бактерій.
Лізоцим — антибактеріальний фактор тваринного походження, що є компонентом неспецифічної резистентності організму. За механізмом дії лізоцим є мурамінідазою. Це гідролітичний фермент, який розщеплює (1→4)-глікозидні зв’язки між дисахаридними компонентами в полісахаридному ланцюгу мураміну, що призводить до руйнування клітинної стінки мікроорганізмів.