Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ IV. МОЛЕКУЛЯРНІ МЕХАНІЗМИ СПАДКОВОСТІ ТА РЕАЛІЗАЦІЇ ГЕНЕТИЧНОЇ ІНФОРМАЦІЇ
ГЛАВА 21. БІОСИНТЕЗ БІЛКІВ У РИБОСОМАХ
21.3. ЕТАПИ ТА МЕХАНІЗМИ ТРАНСЛЯЦІЇ
Молекулярні механізми рибосомальної трансляції у прокаріотів та еукаріотів мають подібні риси і поділяються, як і при синтезі інших біополімерів, на етапи ініціації, елонгації та термінації.
Процес трансляції в клітинах еукаріотів
1. Ініціація трансляції.
Передумовою для початку функціонування рибосомальної білоксинтезуючої системи є утворення ініціюючого комплексу, до складу якого входять:
- субодиниці 40s та 60s, сполучені між собою у SOs-рибосому; цілісна рибосома має дві структурні ділянки для зв’язування в процесі трансляції молекул тРНК, навантажених аміноацильними залишками: аміноацильну (А-) ділянку (А-сайт) та пептидильну (П-) ділянку (П-сайт), перша з яких в ході трансляції є сполученою з аміноацил-тРНК, а друга — з пептидил-тРНК;
- мРНК, що має обов’язково 7-метилгуанозиновий «кеп» на 5'-кінці; мРНК зв’язується з рибосомою таким чином, що напроти її п-ділянки розміщується ініціюючий кодон — AUG, який відповідає, за таблицею генетичного коду, амінокислоті метіоніну — ініціюючій амінокислоті (та взаємодіє з антикодоном мет-тРНК);
- мет-тРНКі — особливий тип тРНК, що акцептує та поставляє в рибосому (спочатку — на П-сайт) першу, ініціюючу, амінокислоту — метіонін (включення метіоніну в середину пептидного ланцюга вимагає присутності іншої тРНК — спеціальної тРНКмет); зв’язування мет-тРНКі з 408-субодиницею рибосом вимагає участі фактора ініціації eIF-2.
Таким чином, метіонін стає N-кінцевою амінокислотою для більшості еукаріотичних білків, його відщеплення з N-кінця можливе на стадії посттрансляційної модифікації пептиду. У прокаріотів першою, ініціюючою, амінокислотою є модифікований метіонін — формілметіонін, що надходить у рибосому у вигляді формілметіоніл-тРНК;
- білкові фактори ініціації (eIF-1, eIF-2, eIF-3 тощо — всього на даний час відомо до десяти факторів ініціації); зокрема, утворення цілісної 808-рибосоми з субодиниць та її стабілізація вимагають присутності факторів ініціації eIF-3, eIF-4С та eIF-6;
- коферменти ГТФ та АТФ, що забезпечують енергією різні етапи ініціації.
Схему будови ініціюючого комплексу подано на рис. 21.4.
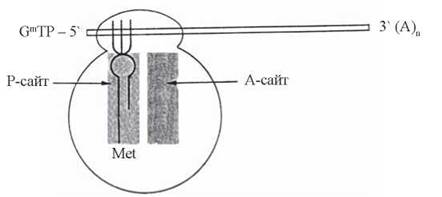
Рис. 21.4. Ініціюючий комплекс, що складається з двох субодиниць рибосоми, м-РНК, мет-тРНК1, білкових факторів ініціації.
GmTP — 7-метилгуанозинтрифосфат (кеп); (A)n — поліаденілатний хвіст мРНК.
2. Елонгація поліпептидного ланцюга.
Суто елонгація полягає в утворенні пептидних зв’язків між амінокислотними залишками, що зв’язані через відповідні тРНК з А- та П-ділянками транслюючої рибосоми. Етапи елонгації схематично подано на рис. 21.5.
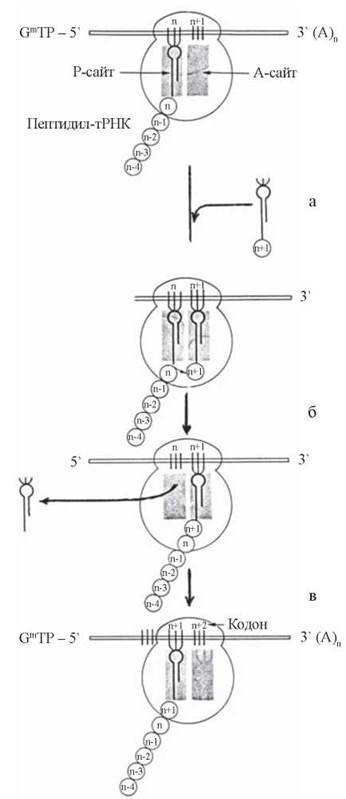
Рис. 21.5. Процес елонгації пептидного ланцюга.
Передумовою початку елонгації є зв’язування з А-сайтом рибосоми (який на даному етапі є вільним) 2-ї (в загальному випадку — (n + 1)-ї, рахуючи з N-кінця пептиду, що синтезується) амінокислоти, сполученої з тРНК (рис. 21.5а). Ця (n + 1)-ша амінокислота відповідає (за генетичним кодом) кодону мРНК, який послідовно йде за ініціюючим (тобто AUG) кодоном.
Пептидилтрансферазна реакція
Утворення пептидного зв’язку між 1-ю (ініціюючою — метіоніном) та 2-ю амінокислотою, що зв’язані через свої тРНК з П- та А-сайтами рибосоми, відповідно) каталізується ферментом пептидилтрансферазою. Пептидилтрансферазна активність пов’язана з 50s-субодиницею прокаріотів та 60s-оубодиницею еукаріотів.
Ця ж пептидилтрансферазна реакція реалізує і подальші етапи елонгації, коли з П- та А-сайтами рибосоми сполучені, відповідно, пептидил-тРНК (містить «n» амінокислотних залишків) та певна наступна («n + 1») амінокислота.
У ході пептидилтрансферазної реакції відбувається перенос пептидного фрагменту (що зв’язаний через відповідну тРНК з П-сайтом) на амінокислоту (що зв’язана через тРНК з А-сайтом) таким чином, що в результаті реакції новий пептид, який утворився, стає зв’язаним з А-сайтом рибосоми. тРНК, що була первинно сполучена з П-сайтом, вивільняється (рис. 21.5б).
Реакція транслокації
Після утворення пептидного зв’язку відбувається переміщення подовженого пептиду, сполученого з тРНК (пептидил-тРНК), з А-сайту в П-сайт — процес транслокації.
Водночас відбувається переміщення рибосоми впродовж ланцюга мРНК вправо. У результаті цього навпроти А-сайта рибосоми розміщується новий (n + 2)-й кодон мРНК, який відповідає наступній — (n + 2)-й амінокислоті, що у вигляді тРНК-комплексу може займати відповідне місце на рибосомі (рис. 21.5в).
У транслокації бере участь білковий фактор елонгації eEF-2. Енергетичні потреби транслокації забезпечуються ГТФ-азною реакцією розщеплення ГТФ до ГДФ.
3. Термінація трансляції.
Термінація трансляції відбувається, коли транслююча рибосома у своєму переміщенні впродовж ланцюга мРНК досягає одного з термінуючих кодонів — UAA, UAG або UGA.
Поява в А-сайті термінуючого кодону розпізнається білковими рилізинг-факторами, які спричиняють гідроліз зв’язку між пептидом та молекулою тРНК, що займає П-сайт рибосоми. У результаті цього процесу відбувається вивільнення пептиду, що синтезувався, та дисоціація 80s-рибосоми на 40s- та 60s-субодиниці.
Посттрансляційна модифікація пептидних ланцюгів
Поліпептидний ланцюг, що є продуктом рибосомальної трансляції, набуває своїх біологічних властивостей після утворення притаманної йому унікальної просторової конформації білкової молекули, чому в багатьох випадках передує його посттрансляційна модифікація (процесинг).
Реакції посттрансляційної модифікації пептидів:
а) модифікація N-та С-кінців — видалення N-кінцевих формілметіоніну (у прокаріотів) та метіоніну (у еукаріотів); ацетилювання N- та С-кінців;
б) модифікація гідроксильних, амінних та карбоксильних груп у бічних радикалах пептидів шляхом їх фосфорилювання, карбоксилювання, метилювання, ацетилювання тощо;
в) приєднання до пептидів простетичних груп — вуглеводів (глікозилювання), гему, коферментів (флавінових нуклеотидів, біотину, порфіринів тощо);
г) хімічна модифікація ковалентної основи амінокислотних залишків; прикладом може бути перетворення у складі фактора ініціації еукаріотів eEF-2 залишку гістидину в залишок незвичайної амінокислоти дифталаміду.