СТРУКТУРА И СВОЙСТВА БИОМОЛЕКУЛ - А. Е. Земляков - 2017
15. ЛИПИДЫ: КЛАССИФИКАЦИЯ И СТРОЕНИЕ
Липиды - низкомолекулярные органические вещества, извлекаемые из клеток неполярными растворителями. Главная биологическая функция липидов заключается в участии в образовании биологических мембран.
По химическому строению липиды очень разнообразны. К ним относят производные высших кислот, спиртов, альдегидов, стероидов. В строении большинства из них можно выделить с одной стороны гидрофильные, полярные группировки, а с другой - гидрофобные (липофильные), неполярные радикалы. В ряде структур полярная и неполярная части связаны через соединительное звено.
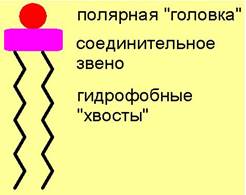
Подобные амфифильные, т. е. обладающие двойным сродством, молекулы легко агрегатизируются в водной среде.
❖ Классификация липидов. Выделяют две основные группы липидов:
✵ Простые (нейтральные) липиды. К ним, в частности, относятся
♦ триглицериды - эфиры глицерина и высших карбоновых кислот;
♦ воски;
♦ стерины.
✵ Сложные (полярные) липиды, которые подразделяются на:
♦ фосфолипиды - в состав их полярной части входит фосфорная кислота и ее производные;
♦ сульфолипиды - в полярной части содержатся остатки серной кислоты, которые обычно введены в углеводный фрагмент;
♦ гликолипиды - в этих соединениях роль полярной головки играют моно- и олигосахариды.
По природе соединительного звена липиды делят на две группы.
♦ Глицеролипиды - в основе структуры лежит многоатомный спирт глицерин.
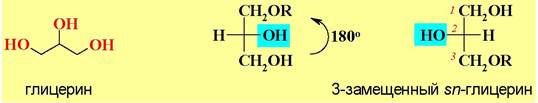
Если сам глицерин оптически неактивен, то введение любого заместителя по первичному гидроксилу делает центральный атом асимметричным. Положение заместителей в молекуле глицерина различают по системе стереоспецифической нумерации (sn). По этой системе, если в проекции Фишера гидроксил у С2 находится слева, то верхний атом получает номер 1.
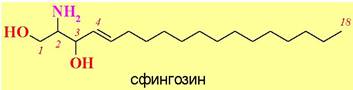
♦ Сфинголипиды - построены на основе липофильного аминоспирта, например, сфингозина.
Простые липиды
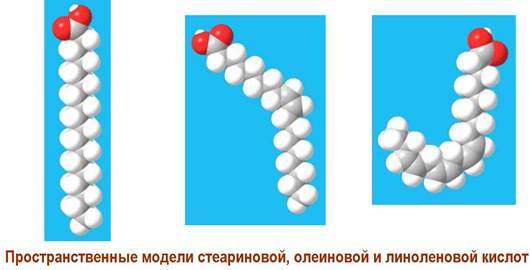
❖ Триглицериды - эфиры глицерина и высших карбоновых кислот. В мембранах растительных и животных клеток присутствуют в незначительных количествах. Являются основным компонентом природных масел и жиров, играющих в организме функцию резерва энергии. Гидроксильные группы глицерина обычно ацилировны различными карбоновыми кислотами. Повышенное содержание ненасыщенных кислот в растительных триглицеридах обеспечивает им жидкое агрегатное состояние (масла), тогда как насыщенные кислоты приводят к твердой консистенции триглицеридов животного происхождения (жиры).
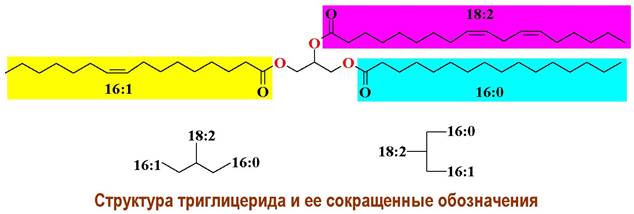
❖ Жирные кислоты. Компонентами всех трех групп липидов являются высшие карбоновые кислоты самого различного строения. В природе обнаружено более 500 жирных кислот. Структуры основных жирных кислот представлены далее в таблице 10. Карбоновые кислоты с относительно небольшим числом углеродных атомов (С4-С10) находятся среди липидов молока. Декановая и лауриновая кислоты в достаточно большом количестве найдены среди липидов масел пальм. Самые распространенные в природе кислоты имеют от 16 до 22 углеродных атомов. Существенно реже встречаются жирные кислоты с числом углеродных атомов более 30. Основные источники их выделения - воски и бактериальные липиды.
Таблица 11. Структура и названия основных природных карбоновых кислот
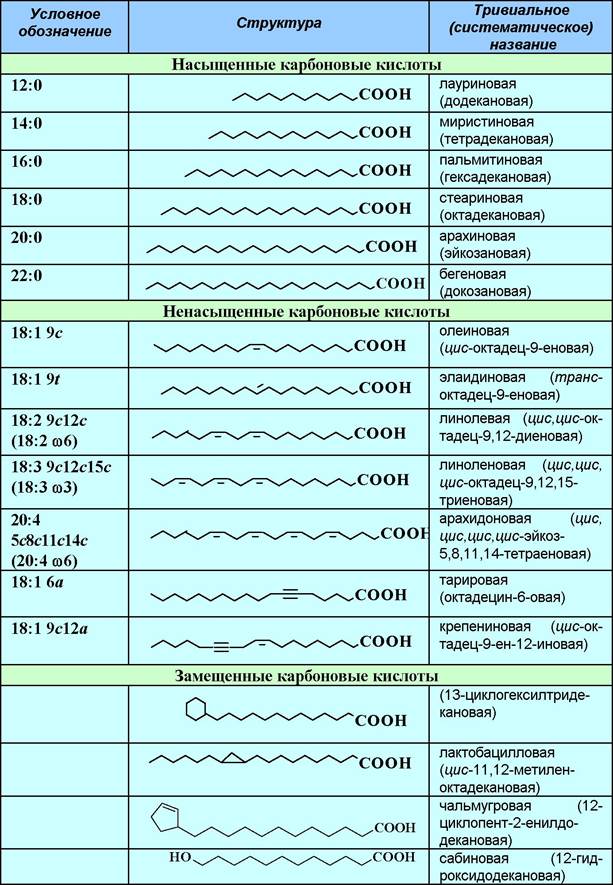
Наиболее распространены насыщенные кислоты с четным числом углеродных атомов и непредельные карбоновые кислоты с одной или несколькими цис-двойными связями. В полиеновых кислотах двойные связи обычно разделены метиленовым звеном. Значительно реже в биологических объектах встречаются кислоты, содержащие тройную связь, цикл (циклогексановый и циклопропановый фрагменты содержат кислоты бактериальных липидов, например, TDM) или гидроксильную группу, например, 3- гидрокситетрадекановая (3-гидроксимиристиновая) кислота в составе липида А.
Для обозначения высших кислот и кислотных остатков используется сокращенная запись, в которой первая цифра обозначает общее количество атомов углерода, а вторая - количество кратных связей; цифры с буквами обозначают локализацию кратных связей: цифра - порядковый номер атома углерода, а буквы - тип и конфигурацию кратной связи (а - тройная связь, с - цис-конфигурация, t - транс-конфигурация). Например, октадец-6-иновая (тарировая) кислота обозначается как 18:1 6а. Данные условные сокращения используют и при записи структур липидов.
Линолевая (18:2 9с12с) и линоленовая (18:3 9с12с15с) кислоты являются незаменимыми кислотами, так как они не синтезируются в организме человека и поступают в него только с пищей. Также очень важна арахидоновая кислота (20:4 5с8с11с14с), которая необходима для биосинтеза таких биологически активных соединений как простагландины, тромбоксаны и лейкотриены. Смесь этих кислот называют витамином F.
Для полиеновых кислот, у которых двойные связи разделены метиленовой группой, используют альтернативный способ обозначения - указывают после буквы ю номер атома углерода первой с конца молекулы двойной связи. Например, линоленовая кислота - 18:3 ω3, арахидоновая кислота - 20:4 ω6.
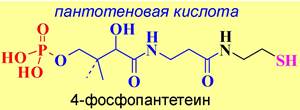
❖ Биосинтез жирных кислот. Синтез пальмитиновой кислоты в клетках животных происходит в полиферментном комплексе - синтазе жирных кислот, представляющей собой димер идентичных блоков по 240 кДа. N- Концевая часть протеина содержит 3 каталитических домена: 3- кетоацилсинтазу (KS), малонил/ацетилтрансферазу (МАТ) и 3- гидроксиацилдегидратазу (DH). В свою очередь, в С-концевом участке выделяют 3 участка с ферментативным действием: еноилредуктазу (ER), 3- кетоацилредуктазу (KR), тиоэстеразу (ТЕ) и ацилпереносящий белок (ACP). Между этими активными участками находится большой (до 600 а. о.) коровый фрагмент. Ацилпереносящий белок содержит в своей структуре остаток 4- фосфопантетеина, тиольная группа которого способна ацилироваться, и обеспечивает последовательное перемещение остатка карбоновой кислоты от одного фермента к другому.
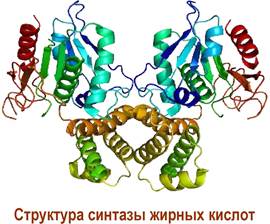
Синтез начинается с переноса ацетильной группы с молекулы ацетил- СоА на ACP при катализе малонил/ацетилтрансферазой. Далее ацетильный фрагмент перемещается на тиольную группу остатка цистеина в KS-домене, а тиольная группа ACP ацилируется малонил-СоА.
Взаимодействие ацетил-KS и малонил-ACP приводит к переносу двух атомов углерода от первой молекулы на вторую с одновременным выделением диоксида углерода. В ацетоацетил-ACP последовательно восстанавливается кетогруппа, гидроксильная группа элиминируется и восстанавливается двойная связь. Цикл превращений завершается бутирил-KS - т. е. исходная углеродная цепь ацетил-KS выросла на два атома углерода.
Новый цикл начинается с взаимодействия бутирил-KS с малонил-ACP, а заканчивается KS-производным капроновой кислоты. Конечным этапом является образование пальмитоил-ACP, который расщепляется под действием гидролитической тиоэстеразы с выделением пальмитиновой кислоты.
Таким образом, в животных клетках конечным продуктом биосинтеза является пальмитиновая кислота. Дальнейшие биохимические трансформации позволяют получить предельные и непредельные кислоты с большим или меньшим числом атомов углерода.
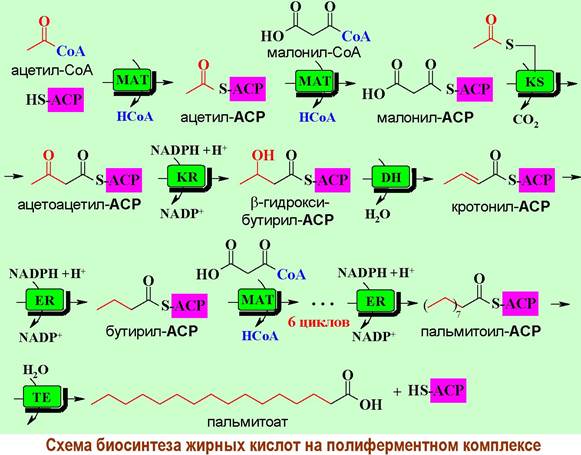
В растительных клетках возможно отделение от ферментативного комплекса меньших по размеру карбоновых кислот. Четное число атомов углерода в этих кислотах связано с последовательным присоединением двухуглеродных фрагментов от малонил-СоА.
❖ транс-Ненасыщенные кислоты. В последние годы в обществе широко обсуждается проблема влияния на организм человека трансненасыщенных кислот. Основным источником этих кислот является маргарин, где их содержание обычно составляет 5-10% от общего количества жирных кислот, а в некоторых образцах достигает 50%. транс-Ненасыщенные кислоты образуются в процессе гидрогенизации остатков цис-непредельных карбоновых кислот растительных масел. На катализаторах наряду с процессами восстановления происходит и изомеризация двойных связей.
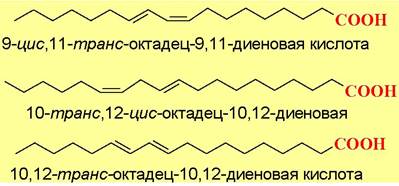
Ниже представлены структуры некоторых транс-кислот маргаринов, из которых основной является 9-цис, 11-транс-октадец-9,11-диеновая кислота.
❖ Воски. В эту группу объединяют продуцируемые животными и растениями липиды, представляющие собой сложные эфиры высших карбоновых кислот и высших спиртов или стеринов. Обычно воски являются смесью эфиров, высших карбоновых кислот и спиртов.
Типичными представителями восков животного происхождения являются:
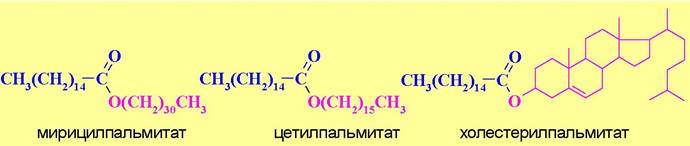
♦ пчелиный воск (главный компонент - мирицилпальмитат),
♦ спермацет кашалотов (основа - цетилпальмитат),
♦ ланолин - воск шерсти овец, в основе которого - холестерилпальмитат).
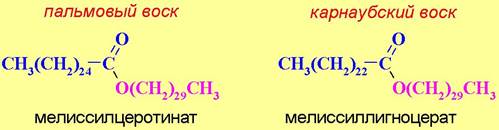
Примерами растительных восков могут послужить пальмовый и карнаубский воски из восковой и бразильской пальм. Основными компонентами этих восков являются эфиры мелиссилового спирта (С30Н61ОН) церотиновой (С25Н51СООН) и лигноцеровой (С23Н47СООН) кислот соответственно.
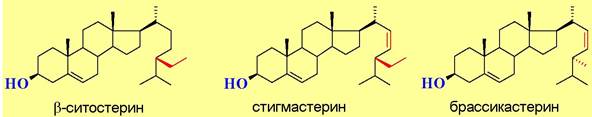
❖ Стерины - неомыляемые липиды, имеющие в основе своего строения циклическую систему эстрана. Выделяют группы:
♦ фитостерины, включающая более 200 соединений, в т. ч. основной стерин высших растений β-ситостерин, а также стигмастерин (соевое масло) и брассикастерин (масло рапса);
♦ микостерины, например, эргостерин, обнаруженный в грибах и дрожжах и являющийся источником синтетического получения витамина D;
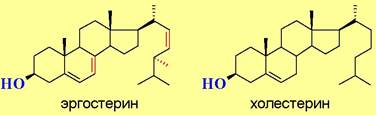
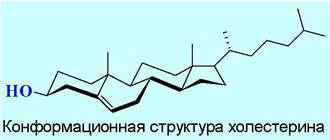
♦ зоостерины, например, холестерин, широко представленный в клетках животных. Мембраны содержат 7-15% стерина. Еще больше (до 25%) холестерина находится в миелиновой оболочке нервных клеток и в мембранах эритроцитов. Наличие холестерина сильно влияет на подвижность и проницаемость липидных слоев. Является предшественником в биосинтезе стероидных гормонов и витамина D. Около 80% холестерина человек получает в результате биосинтеза и лишь около 20% - с пищей. Наряду со стероидными спиртами в природе также часто встречаются сложные эфиры холестерина.
Сложные липиды
❖ Фосфолипиды (ФЛ) - главные компоненты липидных мембран. С точки зрения химического строения это производные эфиров фосфорной кислоты и двух спиртов - глицерина и сфингозина.
✵ Глицерофосфолипиды (ГФЛ).
♦ По природе гидрофобных остатков подразделяют на:
а) диацильные ГФЛ,
б) алкилацильные ГФЛ - содержат остаток высшего спирта; редкие липиды, наиболее распространены в моллюсках;
в) плазмалогены - представляют собой виниловые эфиры, которые при гидролизе образуют высшие альдегиды; содержатся практически во всех организмах.
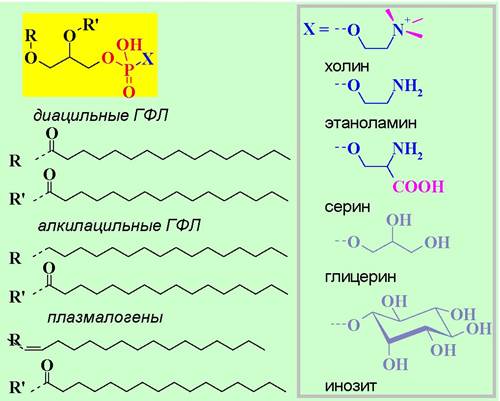
♦ По природе заместителя у остатка фосфорной кислоты различают:
(I) фосфатидилхолин (лецитин), содержащийся в тканях высших растений и животных (до 50% от ФЛ);
(II) фосфатидилэтаноламин (кефалин) - второй по значимости ФЛ, распространен и в бактериальных клетках (15-30%);
(III) фосфатидилсерин - особенно много в головном мозге (до 15% от всех липидов), сердце, почках, селезенке (~10%);
(IV) фосфатидилглицерин - главный ФЛ бактерий (до 70%), много его и в клетках растений (20-30%);
(V) фосфатидилинозит - в качестве полярной компоненты содержит циклический многоатомный спирт миоинозит. Он содержится практически во всех тканях животных (до 10% от всех ФЛ), а также в растениях и микроорганизмах, в частности, входит в структуру липоарабиноманнана клеточных стенок микобактерий.
✵ Сфингофосфолипиды находятся в животных тканях, преимущественно в мембранах клеток мозга и нервной ткани, а также в мембранах эритроцитов (30-40% от общего содержания липидов). Наиболее широко представлены фосфоэфиры N-ацилсфингозинов (церамидов). Из ацильных компонентов обычно встречаются насыщенные и моноеновые кислоты: стеариновая, лигноцериновая (24:0), нервоновая (24:1 15с).
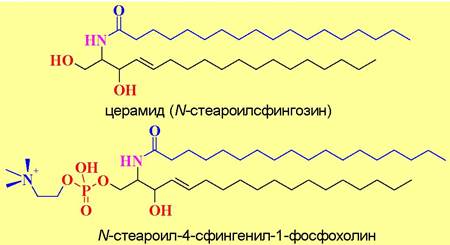
Церамиды не только компоненты биомембран, они играют важную роль в качестве сигнальных молекул в жизненном цикле клеток, в том числе в клеточной дифференцировке, в ингибировании клеточной пролиферации, и стимуляции апоптоза - программированной гибели клеток
❖ Гликолипиды. В качестве полярной части в этой группе липидов выступают моно- или олигосахариды. Углеводный компонент соединен с липидной частью О-гликозидной связью или сложноэфирной связью. К другому типу гликолипидов относится липид А.
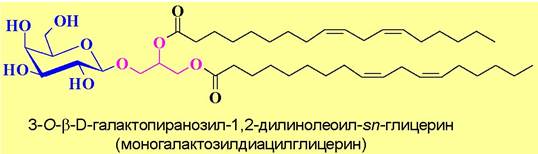
✵ Глицерогликолипиды являются основными компонентами липидов хлоропластов. В этих гликолипидах первичный гидроксил диацилглицерина обычно гликозилирован глюкозой или галактозой.
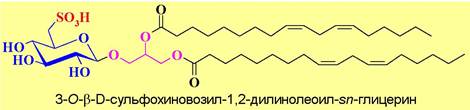
✵ Сульфогликолипиды достаточно широко представлены в клетках растений. Типичным представителем этой группы гликолипидов является сульфохиновозилдиацилглицерин.
✵ Трегалозы димиколат (TDM) - бактериальный гликолипид, представляющий собой диэфир невосстанавливающего дисахарида трегалозы и миколевых кислот.
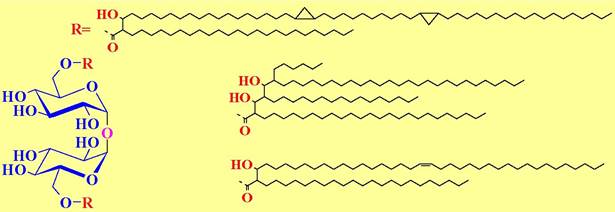
Миколевые кислоты - высшие α-разветвленные β-гидроксикислоты с длиной углеродной цепи от ~30 до более 100 атомов углерода (некоторые из структур миколевых кислот приведены на рисунке). TDM обладает иммуностимулирующим и противоопухолевым действием, однако его применению в медицине препятствует высокая токсичность.
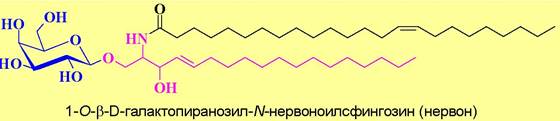
✵ Гликозилцерамиды являются гликозидами N-ацилсфингозина (церамида). Они встречаются в мозге, селезенке, почках, печени. Углеводная часть представлена глюкозой, галактозой, N ацетилглюкозамином, N- ацетилгалактозамином. Выделяют группы:
♦ цереброзиды - углеводная часть моносахаридная (обычно галактоза, реже глюкоза), например, нервон;
♦ глобозиды - церамиды гликозилированы олигосахаридами;
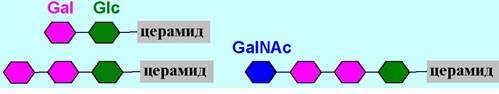
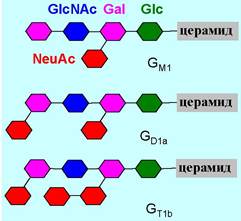
♦ ганглиозиды - в их состав наряду с тетрасахаридным блоком входят сиаловые кислоты, например, N-ацетилнейраминовая кислота (NeuNАс). Особенно много ганглиозидов находится в клетках серого вещества головного мозга. Они в основном расположены на поверхности фосфолипидных мембран и участвуют в процессах роста клеток, межклеточной адгезии, рецепции и др.
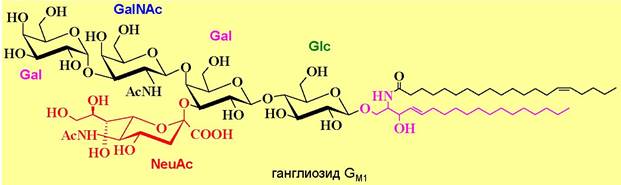
Ганглиозиды обозначают буквой G, буквенными индексами показывают количество остатков нейраминовой кислоты (М - один, D - два, Т - три), остальные цифры и буквы в индексе кодируют расположение этих кислот.