Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Химические основы жизни
Липиды
Жирные кислоты и родственные липиды
Липидами называют соединения биологической природы, растворимые в неполярных растворителях (бензоле, хлороформе, эфире и т. п.) и практически нерастворимые в воде. Из такого определения следует, что липиды могут иметь различное химическое строение и выполнять различные биологические функции. Их относительно низкая растворимость в водных средах является причиной того, что липиды встречаются в основном в неводных биологических фазах, в особенности в клеточных мембранах и мембранах органоидов. К липидам относятся жиры, представляющие собой резервы полимерного биологического топлива, а также некоторые важные медиаторы биологических процессов. Липиды также входят в состав более сложных соединений, например липопротеинов и липосахаридов, которые опять-таки в основном располагаются в биологических мембранах клеток и во внешних оболочках некоторых вирусов.
Насыщенные жирные кислоты представляют собой относительно простые липиды общей формулы СН3(СН2)nСООН. В процессе биосинтеза углеводородная цепь жирных кислот строится из идентичных мономерных звеньев с двумя атомами углерода, поэтому жирные кислоты можно рассматривать как не несущие информации биополимеры с концевой карбоксильной группой. В биологических системах п обычно принимает четные значения от 12 до 20.
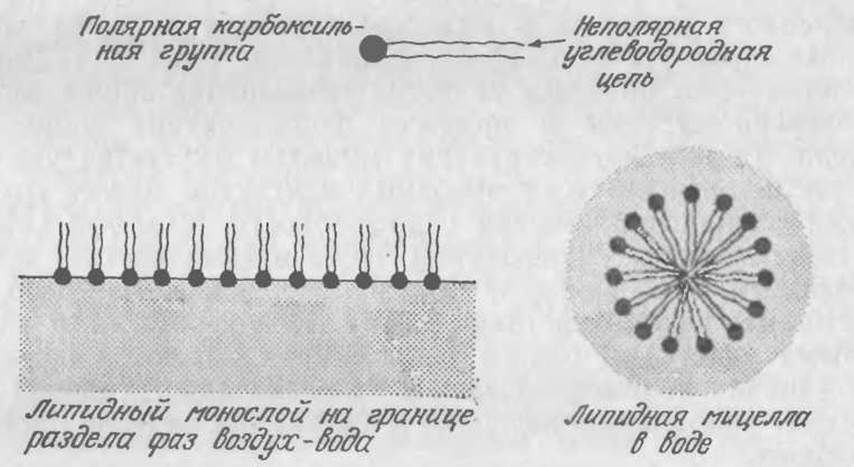
РИС. 2.1. Некоторые устойчивые агрегированные состояния жирных кислот в воде.
Ненасыщенные жирные кислоты образуются при замене насыщенной (—С—С—) связи на двойную связь (—С=С—). Например, олеиновая кислота является ненасыщенным аналогом стеариновой кислоты (n=16):
![]()
Углеводородная цепь придает этим соединениям гидрофобные свойства, но карбоксильная группа в высшей степени гидрофильна. Поэтому, когда жирная кислота находится на границе раздела фаз воздух — вода, небольшое ее количество образует ориентированный мономолекулярный слой (монослой), в котором полярные карбоксильные группы связаны водой, а углеводородные цепи направлены в сторону воздушной фазы (рис. 2.1). Это явление лежит в основе механизма действия моющих средств, представляющих собой соли жирных кислот. Образование мыльного монослоя значительно снижает поверхностное натяжение на границе воздух — вода, что резко повышает способность раствора смачивать и очищать загрязненные места.
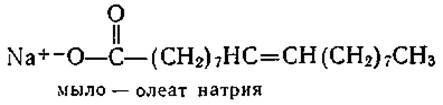
Такого рода гидрофобно-гидрофильные молекулы липидов обладают очень невысокой растворимостью; повышение концентраций раствора выше того предела, который необходим для создания монослоя, приводит к агрегированию избытка растворенного вещества в виде сравнительно больших упорядоченных структур, называемых мицеллами (рис. 2.1). Движущей силой этого процесса является уменьшение общей свободной энергии системы в процессе формирования мицелл из раствора. Та или иная структура мицеллы диктуется увеличением числа энергетически выгодных контактов между группировками одинаковой степени гидрофобности (гидрофильности) и соответствующим уменьшением числа взаимодействий между гидрофобными и гидрофильными группировками. Известно, что аналогичные взаимодействия между гидрофобными и гидрофильными участками одного биополимера являются причиной существования полимерной цепи в одной предпочтительной конформации. Такое поведение ДНК и белков мы вкратце рассмотрим позднее.
Жиры, выполняющие важную функцию внутриклеточного топлива, представляют собой сложные эфиры, образующиеся при конденсации жирных кислот с глицерином:
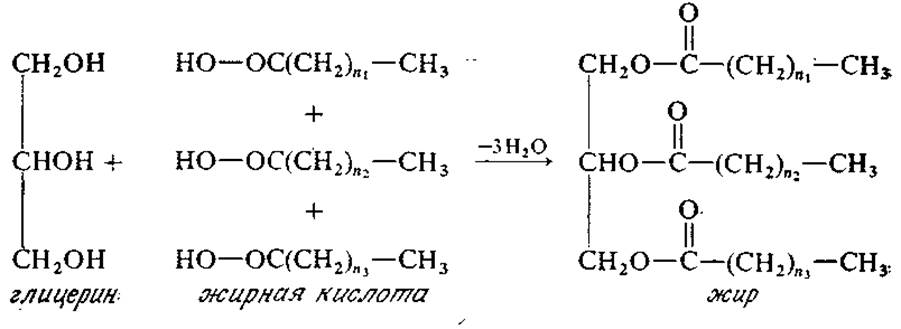
В щелочной среде при нагревании жиры и другие липиды, рассматриваемые в настоящем разделе, гидролизуются до глицерина и солей жирных кислот (мыла) — именно таким путем мыла были впервые получены из животных жиров. Подобная реакция, обратная приведенной выше схеме синтеза жиров, в пищеварительном тракте животных осуществляется при температуре тела и катализируется особыми ферментами, способными расщеплять жиры; микроорганизмы также продуцируют такие ферменты, роль которых заключается в гидролизе некоторых жиров на более мелкие фрагменты, способные затем проникать в клетку через клеточные мембраны.
По строению (но не по выполняемой ими функции) жирам близки фосфоглицериды. В молекулах последних остаток фосфорной кислоты замещает один из концевых остатков жирной кислоты. В результате получается соединение, в молекуле которого опять-таки имеются гидрофильные и гидрофобные остатки, поэтому фосфоглицериды в достаточно высоких концентрациях также способны образовывать мицеллы. Если в раствор фосфолипида поместить пластинку с небольшим отверстием, то в последнем может сформироваться плоский бимолекулярный слой (бислой) толщиной около 70 Å (7∙10-7 см) (рис. 2.2, а). Биологические плазматические мембраны обычно содержат значительные количества фосфолипидов и других полярных липидов. Кроме того, для плазматических мембран характерен хорошо различимый на электронных микрофотографиях бимолекулярный слой (рис. 2.2,6) примерно такой же толщины, как и изображенный на рис. 2.2, а самопроизвольно образующийся двойной фосфоглицеридный слой. По этой причине синтетические бислойные липидные мембраны могут служить удобной моделью для изучения основных особенностей тонких биологических мембран.
По ряду физических свойств липидные бислойные мембраны напоминают клеточные мембраны. Оба типа мембран имеют высокое электрическое сопротивление и значительную электрическую емкость, что в большой степени обусловливает непроницаемость природных мембран для веществ, несущих электрический заряд, например фосфорилированных соединений. Это свойство мембран в свою очередь позволяет клеткам хранить запас заряженных питательных веществ и промежуточных продуктов метаболизма, а также поддерживать высокую разность концентраций небольших катионов (Н+, К+, Na+ и т. п.) внутри клетки и вне ее.
Другие компоненты мембран и их роль в обмене веществ между клеткой и средой будут рассмотрены в разд. 2.5 и 5.7. Барьеры и каналы транспорта биохимически важных соединений определяют, какие вещества из числа участвующих в сложной сети катализируемых ферментами процессов проникнут в клетку, какие останутся в ней, а какие будут выведены из клетки. Функции регулирования транспорта веществ обязательны для нормальной жизнедеятельности любой клетки; они также играют важную роль в технологических процессах, в которых клетки являются катализаторами.
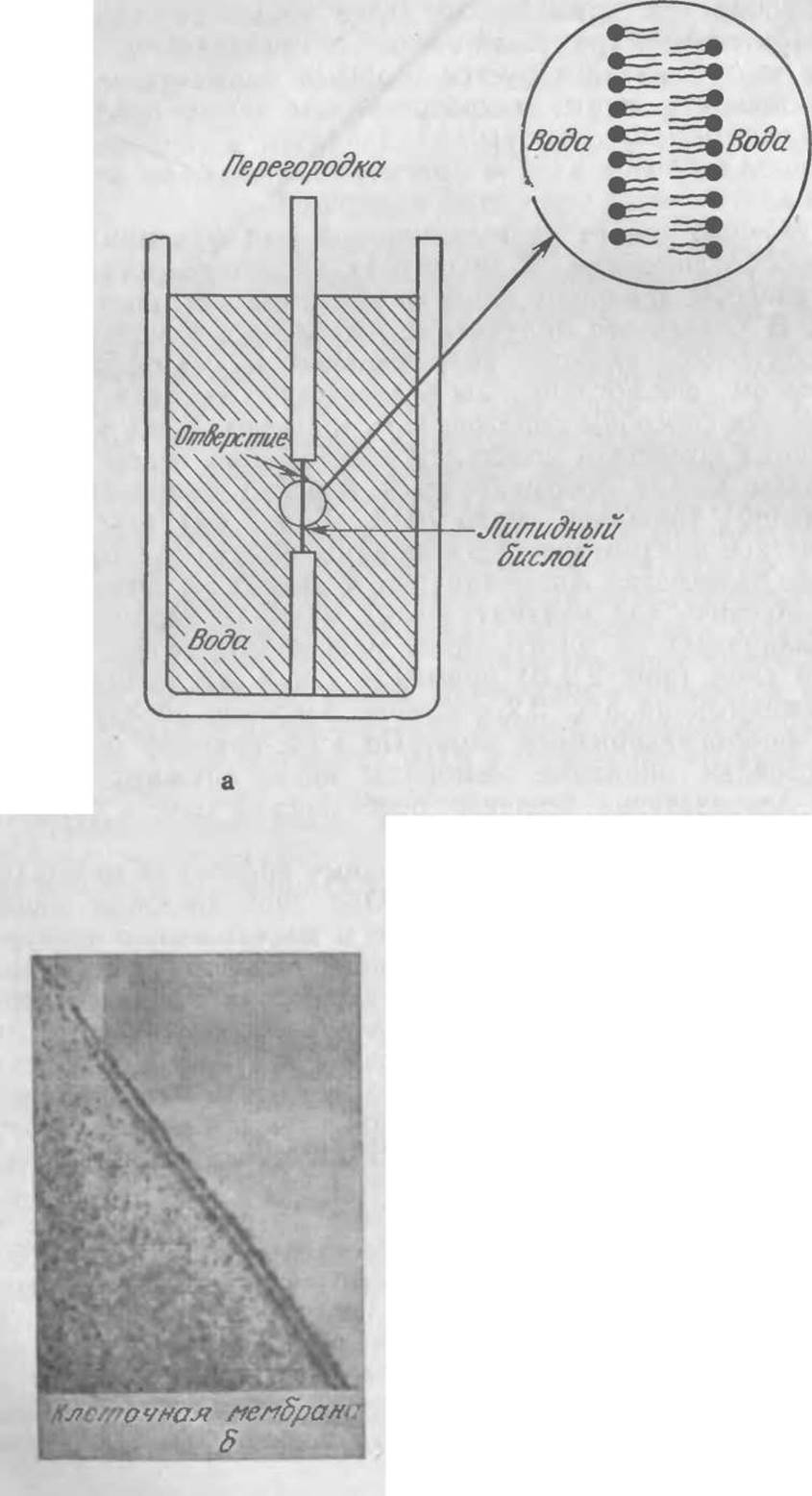
РИС. 2.2. а — самопроизвольное образование устойчивого фосфоглицеридного бислоя в отверстии перегородки, помещенной в сосуд с раствором липида; б — по внешнему виду самопроизвольно образующийся двойной слой напоминает аналогичные структуры клеточных мембран, хорошо различимые на электронных микрофотографиях. [Рис. 2.2,6 воспроизведен с разрешения из работы: Robertson J. В., Membrane Models: Theoretical and Real, In The Nervous System, vol 1: The Basic Neurosciences, Tower B. D. (ed.), p. 43, Raven Press, New York.]
Путем добавления небольшого количества различных веществ можно селективно изменять проницаемость как модельных бимолекулярных, так и плазматических мембран относительно определенных ионов. В частности, было установлено, что некоторые антибиотики и другие соединения, связывающие катионы в комплексы, способны значительно усиливать пассивный ионный транспорт в обеих типах мембран. Более сложные процессы обусловливают повышение проницаемости клеточных стенок живых организмов после их мягкой химической или тепловой обработки. Это свойство клеток с успехом используется в процессах микробиологического производства веществ, являющихся промежуточными соединениями в клеточном метаболизме, а также в целях снижения содержания нуклеиновых кислот в клеточной массе, применяемой в качестве корма для животных.