Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Кинетика процессов утилизации субстрата, образования продуктов метаболизма и биомассы в культурах клеток
Кинетика тепловой гибели клеток и спор
Активность клеток, спор и вирусов в воздухе или жидких средах может снижаться в результате их разрушения (под воздействием тепла, радиации, химических агентов или механических сил), при механическом отделении (фильтрованием или центрифугированием) или за счет ингибирования (при переохлаждении, высушивании, обезвоживании, воздействии химических реагентов). Жидкости стерилизуют в основном путем нагревания (в промышленности) или хлорирования (для бытовых нужд), а для стерилизации воздуха применяют главным образом фильтрование. В двух последних процессах большую роль играют эффекты массопередачи, рассматриваемые в следующей главе; этот раздел посвящен кинетике инактивации клеток и спор.
На рис. 7.33 изображены результаты экспериментального изучения воздействия повышенной температуры на вегетативные клетки и споры. В общем случае по сравнению со спорами клетки погибают при нагревании гораздо быстрее. Это и понятно, если учесть, что образование эндоспор, как мы уже отмечали ранее, представляет собой защитный механизм, позволяющий некоторым клеткам переносить неблагоприятные условия. Тепловая обработка приводит также к инактивации вирусов и бактериофагов, поэтому использующаяся в микробиологической промышленности тепловая стерилизация снижает как численность популяции жизнеспособных микроорганизмов, так и концентрацию (титр) вирусов в подаваемом в реактор потоке питательных веществ.
Прежде чем перейти к изучению уравнений, описывающих скорость снижения численности популяции в процессе стерилизации, необходимо сделать ряд замечаний. Гибель каждой конкретной клетки, по-видимому, обусловливается термической денатурацией одного или нескольких жизненно важных для этой клетки белков, например ферментов. Кинетика подобных кооперативных превращений столь сложных молекул может изменяться во времени самым неожиданным образом. К тому же скорость молекулярных процессов, приводящих в конце концов к гибели клетки, зависит от состава среды, в том числе и от концентрации растворителя. Например, как показано в табл. 7.9, температура, при которой наступает коагуляция (денатурация с последующей интенсивной необратимой сшивкой молекул денатурированного белка) альбумина яичного белка, повышается с уменьшением содержания в системе воды.
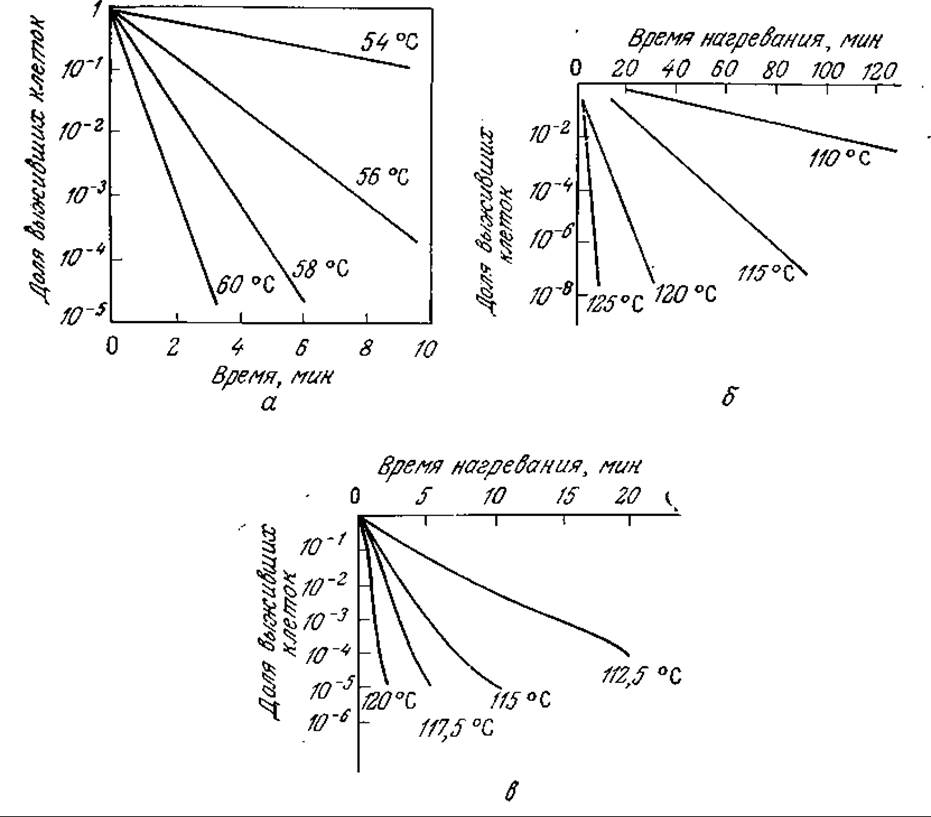
РИС. 7.33. Результаты экспериментального определения скорости тепловой гибели Е. coli и тепловой инактивации вегетативных спор Bacillus subtilis и вегетативных спор В. słearothermophilus. [Воспроизведено из работ: а) Аиба Ш., Хемфри А., Миллис Н., Биохимическая технология и аппаратура.— М.: Пищевая промышленность, 1975; б, в) Burton Н., Jayne-Williams D., Sterilized Milk, in Recent Advances in Food. Science, vol. 2: Processing, Hawthorn J., Leitch J. M. (eds.), p. 107, Butterworths & Co., Publishers Ltd., London, 1962.] a — скорость гибели E. coli в буферном растворе; б — скорость инактивации спор Bacillus subtilis; в — скорость инактивации спор Bacillus stearothermophilus.
Эти данные и ряд других экспериментально обнаруженных фактов свидетельствуют о том, что в случае сравнительно обезвоженных организмов и структур (например, вирусов или спор) их инактивацию правильнее рассматривать как двухэтапный процесс, при котором сначала происходит гидратация, а затем, возможно, и денатурация. Такому механизму отвечает зависимость скорости гибели от относительной влажности, обнаруженная в случае бактериофагов [34]. Совместное влияние нескольких инактивирующих факторов не обязательно носит аддитивный характер; так, совместные обезвоживание и тепловая обработка могут быть менее эффективными, чем можно было бы предположить, судя по эффективности каждого из этих факторов в отдельности.
Таблица 7.9. Зависимость температуры коагуляции альбумина от содержания водыа
Содержание воды, % |
Ориентировочная температура коагуляции, °С |
Содержание воды, % |
Ориентировочная температура коагуляции, °С |
50 |
56 |
5 |
149 |
25 |
76 |
0 |
165 |
15 |
96 |
а Данные из работы: Frobisher М., Fundamentals of Microbiology, р. 259, W. В. Saunders Company, Phila., 1968.
Как мы упоминали выше, популяция состоит из множества клеток различного возраста. Природа клеточной стенки и относительная важность любого из путей метаболизма зависят от возраста, данной клетки и культуры в целом. Следовательно, устойчивость клеток к тепловой или индуцированной иными факторами инактивации будет зависеть от их истории, т. е. от довольно неопределенного фактора, который далеко не всегда удается охарактеризовать количественно. В частности, клетки популяции, находящейся в фазе экспоненциального роста, имеют относительно проницаемые стенки, что способствует эффективному обмену растворенными веществами между внутриклеточным объемом и средой. Если эти растворенные вещества влияют на устойчивость белков, то можно ожидать, что их присутствие окажет влияние и на рост клеток в экспоненциальной фазе. Поэтому наиболее надежные данные о поведении популяции могут быть получены только в результате экспериментов в условиях, максимально приближенных к условиям предполагаемого практического проведения процесса.
Анализ кинетики скорости гибели клеток мы начнем с замечания о том, что линейная зависимость логарифма доли оставшихся в живых клеток от времени (рис. 7.33) указывает на первый порядок процесса снижения численности популяции жизнеспособных клеток n:
![]()
Отсюда следует, что при постоянном kd
![]()
Здесь n0 — концентрация спор или вегетативных клеток при t = 0. Наклон прямой полулогарифмической зависимости равен — kd; график зависимости lnkd от обратной температуры также представляет собой прямую, поэтому зависимость kd от температуры может быть выражена уравнением аррениусовского типа:
![]()
Для многих спор и вегетативных клеток величина параметра Ed лежит в диапазоне от 50 до 100 ккал/моль. Другой ранее применявшийся параметр Dr, равный 2,303/kd и называемый временем децимальной редукции (десятичным коэффициентом снижения численности популяции), представляет собой не что иное, как время, в течение которого численность популяции жизнеспособных клеток снижается в 10 раз.
Приведенные выше кинетические уравнения справедливы только при том условии, что количество спор или клеток в популяции достаточно велико с точки зрения статистики (разд. 7.1). По мере снижения числа клеток или спор (n) вероятность отклонений от величин, предсказываемых указанными уравнениями, возрастает, поскольку, например, при нормальном распределении стандартное отклонение возрастает пропорционально 1/n. Из предыдущего уравнения детерминистической модели следует, что оставшаяся жизнеспособной доля популяции (если допустить, что в данных летальных условиях роста клеток не происходит) определяется уравнением
![]()
Если судьба каждого организма не зависит от других организмов, если организмы не самовоспроизводятся и если летальные условия одинаковы для каждого организма, то стохастический анализ процесса стерилизации показывает, что вероятность наличия в популяции N жизнеспособных организмов в любой момент времени t равна [33]
![]()
Здесь N0 — численность жизнеспособных организмов в стерилизуемой жидкости непосредственно, перед началом стерилизации.
В этом уравнении параметр kd имеет тот же смысл, что и константа скорости в уравнении (7.124); в стохастической модели kd можно интерпретировать как величину, обратную средней продолжительности жизни организма. По мере того как число организмов снижается до некоторой сравнительно малой величины, допущение о гомогенной популяции, достаточно полно описываемой одним параметром kd, становится все менее и менее обоснованным. Так, приведенные на рис. 7.34 данные, характеризующие поведение Staphylococcus aureus в нейтральном фосфатном буфере, свидетельствуют об отклонении от поведения, предсказываемого стохастическим распределением, в положительную сторону; следовательно, большей устойчивостью к тепловой обработке отличается небольшая доля популяции.
Иногда допускается, чтобы после стерилизации оставалось довольно значительное количество жизнеспособных организмов (так, в пастеризованном молоке высшего качества концентрация живых бактерий не должна превышать 30 000 клеток в 1 мл). В других ситуациях, например при выращивании чистых культур, к стерилизации предъявляются значительно более жесткие требования. В таких случаях важно оценить вероятность гибели популяции, т. е. вероятность инактивации всех организмов. Подставляя в уравнение (7.128) N = 0, получаем
![]()
Отсюда следует, что вероятность выживания по меньшей, мере одного организма составляет
![]()
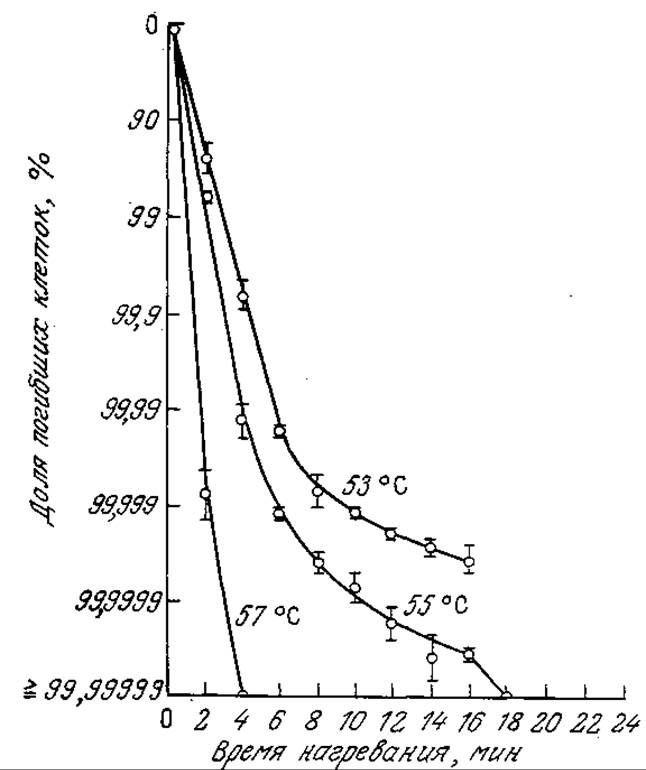
РИС. 7.34. Тепловая инактивация 5. aureus. [Воспроизведено с разрешения из статьи: Walker G. С., Harmon L. G., Thermal Resistance of Staphylococcus aureus in Milk, Whey, and Phosphate Buffer, Appl. Microbiol., 14, 584 (1966).]
Обычно N0 ≫ 1, поэтому
![]()
где
![]()
Величину (1—P0) можно интерпретировать как долю стерилизаций, которые не должны приводить к полной гибели всех организмов.
Изучив метаболизм клетки, мы узнали, что живые клетки часто располагают несколькими альтернативными путями усвоения энергии и осуществления биосинтеза. Наличие таких резервных систем повышает шансы организма на выживание в неблагоприятных условиях. Статистический анализ показывает, что с учетом существования резервных систем численность популяции должна составлять
![]()
где ω — число типов жизненно важных субклеточных структур; r — число единиц каждой структуры в каждом организме; ![]() — средняя удельная скорость разрушения структуры каждого типа.
— средняя удельная скорость разрушения структуры каждого типа.
Обратите внимание на то, что это уравнение можно выразить через ряд убывающих экспоненциальных функций и, следовательно, в полулогарифмических координатах большие участки соответствующей графической зависимости не должны представлять собой прямую линию.
Самый простой подход к анализу популяции из т различных типов клеток (спор или вирусов) также сводится к допущению о независимости видов организмов; тогда суммарная популяция жизнеспособных организмов будет равна сумме индивидуальных популяций, выражаемых уравнениями типа (7.125) с соответствующими индивидуальными коэффициентами kdi:
![]()
И снова график этой зависимости в полулогарифмических координатах не должен представлять собой прямую. Далее, значения индивидуальных параметров kdi не всегда известны; некоторые методы оценки верхнего и нижнего пределов значений n(t)/n0 по начальным данным обсуждены Хатчинсоном и Лассом*. Достоверность таких оценок зависит от точности исходных данных о гибели популяции и требует определения второй производной d2n/dt2 при t = 0.