Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Процессы переноса в биотехнологических системах
Неньютоновские жидкости
Потребляемая мощность и массообмен в биотехнологических процессах с неньютоновскими жидкостями
Мгновенное значение числа Рейнольдса Rei' мешалки, согласно Калдербэнку [11], можно определить по уравнению
![]()
Здесь Кс — индекс вязкости, равный напряжению сдвига т при скорости сдвига 1 с-1.
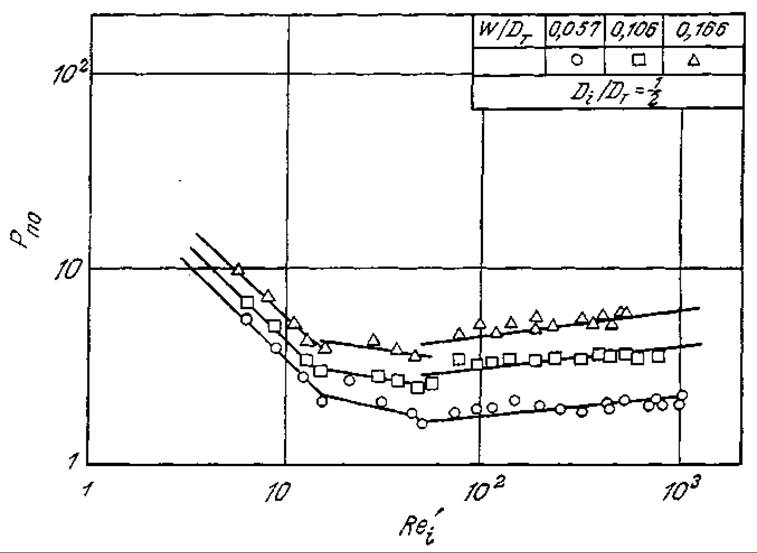
РИС. 8.13. Зависимость числа мощности от числа Рейнольдса Rei (см. текст) в неаэрируемых системах для трех различных типов мешалок. [Из статьи: Taguchi Н., Miyamoto S., Power Requirements in Non-Newtonian Fermentation Broth, Biotech. Bioeng., 8, 43 (1966).]
Как показано на рис. 8.13, для неаэрируемой, неньютоновской культуральной жидкости Endomyces число мощности [уравнение (8.78)] зависит от числа Рейнольдса Rei'. Эту зависимость можно выразить уравнением
![]()
где Di — диаметр лопастной мешалки; DT — диаметр реактора; W — ширина лопасти мешалки; k, х, у и z зависят от диапазона Rei' следующим образом:
Rei' |
|||
<10 |
10-50 |
>50 |
|
k |
32 |
11 |
9 |
x |
-0,9 |
-0,4 |
-0,05 |
у |
-1,7 |
-1,7 |
-1,2 |
z |
0,4 |
0,5 |
0,9 |
Обращает на себя внимание сходство кривых, приведенных на рис. 8.13, с данными Раштона и с кривыми зависимости коэффициента трения в трубе от числа Рейнольдса (рис. 8.8,а,б).
Этими же исследователями показано, что в случае процессов с аэрацией турбулентному режиму при Rei' > 50 удовлетворяет уравнение Мичела и Миллера, применявшееся ранее для ньютоновских жидкостей:
![]()
Оказалось также, что ламинарный и второй режимы (Rei' < 50) с удовлетворительной степенью приближения можно описать тем же выражением с меньшим показателем степени:
![]()
Две кривые пересекаются при P2NiDi3/Fg0,56 = 2⋅10-2; коэффициент пропорциональности в небольшой степени зависит от конструкции перемешивающего устройства.
Разведение мицелиальных культур небольшим количеством (10—25%) воды заметно изменяет klи число мощности. Отсюда следует, что в непрерывных биологических процессах скорость разведения может в существенной степени изменять величину потребляемой мощности.
Влияние популяции микроорганизмов зависит от ее физического состояния (отдельные суспендированные клетки, скопления клеток, клетки, локализованные вблизи поверхности частиц или пузырьков и т. д.) и характера воздействия микроорганизмов на свойства системы. Показано, что при повышении концентрации биомассы в мицелиальной культуре Aspergillus niger от 0,02 до 2,5% kla снижается на 90%. С другой стороны, наблюдалось повышение скорости массообмена О2 в присутствии суспензий Candida intermedia, Pseudomonas ovalis или частиц оксида алюминия диаметром 0,3 мкм. С помощью ингибиторов окислительного фосфорилирования (гл. 5) установлено, что сходство эффектов клеток и оксида алюминия не ограничивается качественными характеристиками; оба фактора повышают скорость массообмена примерно на 40% (по сравнению с водой), причем вызываемый клетками эффект не зависит от их жизнеспособности. Недавно было показано, что влияние этих агентов объясняется изменением гидродинамических условий вблизи границы раздела газ — жидкость, сопровождающимся снижением сопротивления массообмену в жидкой пленке, окружающей пузырьки газа [44].
При высоких скоростях сдвига, необходимых для смешения многофазных систем и обеспечения необходимых скоростей массообмена, часто наблюдается снижение активности микроорганизмов. Так, серьезное снижение жизнеспособности популяции относительно большой клетки, простейшего Tetrahymеnа, при механическом повреждении клеток начинает проявляться при скорости сдвига выше 1200 с-1. В этих экспериментах максимальная скорость сдвига (характеризуемая окружной скоростью мешалки) оказалась более важной переменной, чем число Рейнольдса или мощность, потребляемая единицей объема реактора.
Влияние механического перемешивания может проявляться и иным образом; выше мы уже упоминали, что при перемешивании может снижаться размер существующих микробных скоплений. Тагучи и Иошида* рекомендовали разделять экспериментально наблюдаемое снижение размеров гранулы мицелия на два процесса: 1) отщепление значительно меньших частиц от больших гранул; 2) непосредственное разрушение сферической формы гранул. Оказалось, что обусловленное первым процессом уменьшение диаметра D частиц во времени подчиняется закону
![]()
Второй процесс можно описать уравнением первого порядка:
![]()
Здесь Np — число неразрушенных частиц. Зависимость коэффициента скорости разрушения частиц kr от D, Niи Diможно выразить таким уравнением;
![]()
Тагучи и др. предположили, что последнее уравнение можно согласовать с теоретическими данными, если предположить, что разрыв гранул определяется не столько вязкостным, сколько турбулентным напряжением и что устойчивость гранул непосредственно связана с их прочностью на разрыв, определяемой экспериментально.
Что касается скорости переноса кислорода в суспензиях целлюлозной пульпы, то здесь, как было показано, важны конструкции перемешивающего устройства и реактора; эти данные согласуются с результатами изучения аэрируемых и неаэрируемых суспензий Endomyces, опубликованными ранее Тагучи и Миямото [42]. В 1,6%-ной суспензии пульпы произведение kia описывается уравнением
![]()
где t — характерное время выравнивания концентраций в реакторе; h — высота слоя жидкой фазы; W — ширина лопасти мешалки. Физический смысл параметров Di, DT, L указан выше.
* Taguchi H., Yoshida T., J. Ferment. Technol., 46, 814 (1968).
Подводя итог, можно отметить, что в общем случае коэффициенты массопередачи и величина площади поверхности раздела фаз в единице объема определяются целым рядом факторов, в том числе размерами пузырьков и клеток, реологическими и другими свойствами жидкой фазы (и газожидкостной системы), конструкцией перемешивающего устройства и реактора, потребляемой мощностью. При проектировании и расчете биореакторов необходимо принимать во внимание как все перечисленные факторы, так и параметры кинетики клеточного роста (рассмотренные в предыдущей главе), а также параметры процесса смешения в реакторе и тип последнего (гл. 9).
Очевидно, желательно добиться такого уровня знаний, когда можно было бы охватить все эти вопросы; в то же время надо отдать себе отчет в том, что все изложенные в этом разделе теории и соответствующие уравнения представляют собой всего лишь введение в обширную область знаний, необходимую для проектирования биореакторов на основе одних лишь теоретических разработок. В заключение мы рассмотрим пример, иллюстрирующий некоторые сложные зависимости, природу которых еще предстоит выяснить. В этом примере рассмотрено изменение во времени соотношений между параметрами воздушных пузырьков, диспергированного субстрата и клеток в процессе роста Candida petrophilum на н-гексадекане (рис. 8.14). Заслуживает внимания описание этой своеобразной системы газ — жидкость — жидкость — клетка, данное авторами работы*:
«В первом периоде процесса маслянистые капли относительно велики, и клетки концентрируются в основном вблизи этих капель, а не воздушных пузырьков. Воздушные пузырьки неустойчивы и легко обновляются. Значение kla можно поддерживать на максимальном уровне, зависящем только от конструкции реактора. В течение второго периода маслянистые капли уменьшаются в размере, а клетки адсорбируются на их поверхности, образуя плотные хлопья. Для хлопьев характерна тенденция к связыванию с поверхностью воздушных пузырьков, но перемешивание легко разрушает эти образования.
* Mimura A., Takeda I., Wakasa R., Some Characteristic Phenomena of Oxygen Transfer in Hydrocarbon Fermentation, Biotech. Bioeng. Symp. 4, pt. 1, 467 (1973).
По мере развития процесса величина kla постоянно снижается. Третий период процесса одновременно является вторым полупериодом фазы логарифмического роста, в котором дрожжи продолжают быстро расти, хотя под микроскопом уже не видны маслянистые капельки, В этот момент kla достигает минимального в течение всего процесса значения... [Воздушные] пузырьки покрываются слоем дрожжевых клеток и сближаются, образуя скопления, в которых отдельные пузырьки разделены слоями биомассы. Подобные образования, плавающие на поверхности культуральной жидкости, очень устойчивы. В четвертом периоде н-парафин полностью истощается. Клетки распределяются равномерно по всему объему культуральной жидкости. В этом периоде культуральная жидкость по своим свойствам, вероятно, близка культуральной жидкости, образующейся при брожении углеводов, a kla возрастает до начального уровня».
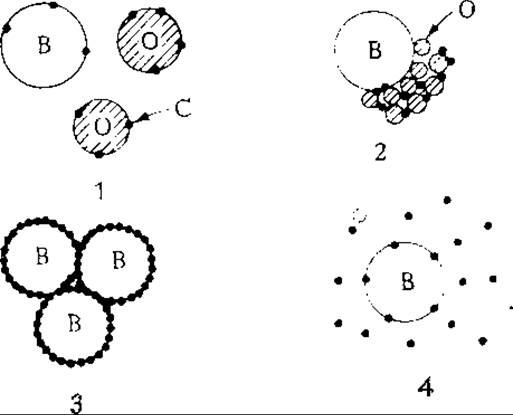
РИС. 8.14. Виды взаимодействия между воздушными пузырьками (В), капельками н-гексадекана (О) и дрожжевыми клетками (С) на различных стадиях периодического роста культуры дрожжей Candida petrophilum (1 — лаг-фаза; 2 — первая половина экспоненциальной фазы; 3 — вторая половина экспоненциальной фазы; 4 — после истощения запасов н-гексадекана). [Из статьи: Mimura A., Takeda I., Wakasa R., Some Characteristic Phenomena of Oxygen Transfer in Hydrocarbon Fermentation, in Adv. Microbial Eng., Siky- ta B., Prokop A., Novak M. (eds.), part 7, p. 467, Wiley-Interscience, New York, 1973.]
Математическая модель, учитывающая ряд особенностей процессов такого типа, будет рассмотрена в гл. 9 (пример 9.2).