Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Стехиометрия и энергетика метаболических превращений
Биосинтез
Синтез макромолекулярных соединений
Как показано на общей карте путей метаболизма, приведенной во введении к настоящей главе, из мономерных веществ-предшественников далее должны строиться полимерные компоненты клетки. Для этой цели опять-таки требуется большое количество метаболической энергии, поскольку любая реакция конденсации сопровождается повышением свободной энергии. Более того, чрезвычайно большие запасы свободной энергии являются той движущей силой, которая резко смещает общее равновесие в сторону образования необходимых полимеров. Это особенно важно в случае биосинтеза полимеров строго определенного строения, например нуклеиновых кислот и белков с заданными последовательностями.
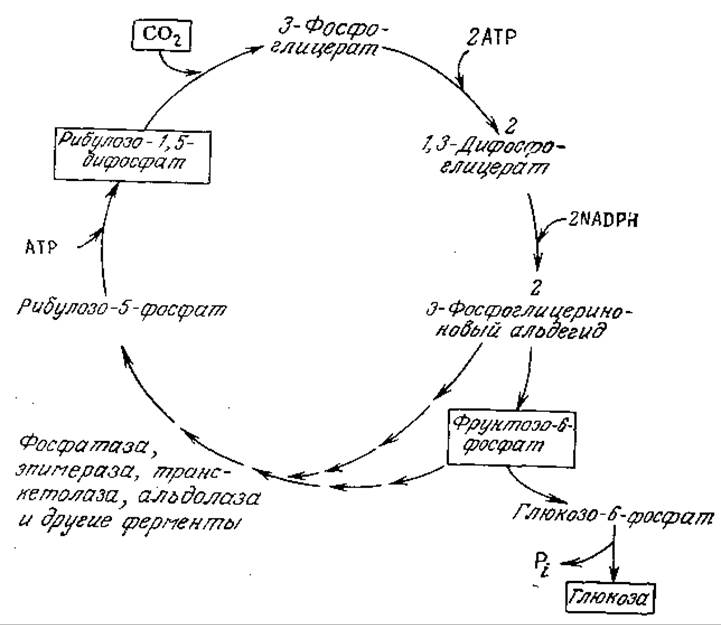
РИС. 5.15. Упрощенная схема синтеза глюкозы из СО2 в цикле Калвина.
На рис. 5.16 показано, что хранящаяся в фосфатных связях АТР энергия при посредстве трифосфатов других нуклеотидов утилизируется в биосинтезе четырех классов биополимеров. Для биосинтеза макромолекулярных соединений типично сочетание реакции конденсации с гидролизом двух высокоэнергетических фосфатных связей, причем сначала нуклеозидтрифосфат превращается в нуклеозидмонофосфат и пирофосфат, который также подвергается гидролизу. Таким путем высвобождается примерно в 2 раза больше свободной энергии (около 14 ккал/моль), чем при превращении нуклеозидтрифосфата в соответствующий дифосфат. Та или иная модификация этого общего механизма реализуется в биосинтезе липидов, РНК, ДНК и гликогена.
Здесь опять-таки используется принцип общих промежуточных соединений, хотя и в несколько усложненной форме. Так, процесс присоединения остатка глюкозы к растущей молекуле гликогена осуществляется в шесть стадий, четыре из которых являются последовательными и, следовательно, связаны общими промежуточными соединениями (табл. 5.5).
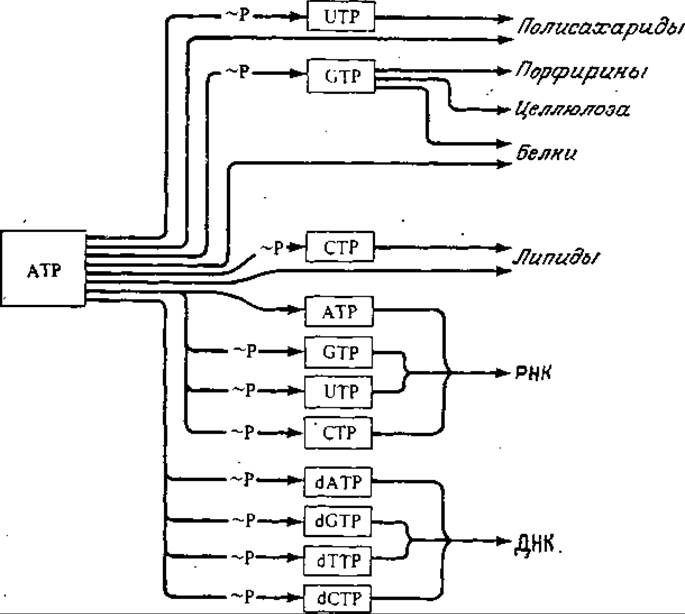
РИС. 5.16. Участие высокоэнергетических трифосфатов нуклеозидов в различных путях биосинтеза. (Воспроизведено с разрешения из работы: Lehninger А., Bioenergetics, 2d ed., р. 136, W. A. Benjamin, Inc., Palo Alto, 1975.)
Естественно, синтез информационных полимеров (РНК, ДНК и белков) представляет собой гораздо более сложный процесс. И в том и в другом случае, однако, наращиванию полимерной цепи предшествует активация мономера. В схему синтеза РНК и ДНК нуклеотиды входят в виде нуклеозидтрифосфатов, а присоединяется к соответствующей полимерной цепи монофосфат; реакция сопровождается образованием пирофосфата. Таким образом, для присоединения одного мономерного остатка необходимо затратить 14 ккал/моль энергии. Активация аминокислот в ходе синтеза белков осуществляется в соответствии со следующей схемой:
![]()
Кроме того, процесс присоединения аденилата аминокислоты к пептидной цепи сопровождается гидролизом GTP; следовательно, для присоединения одного аминокислотного остатка к белковой цепи необходимо гидролизовать в общей сложности три высокоэнергетические фосфатные связи. В гл. 6 мы рассмотрим механизмы биосинтеза заданных последовательностей мономерных остатков в ДНК, РНК и белках.
Таблица 5.5. Последовательные стадии биосинтеза гликогена в клетках млекопитающих (в бактериях и растениях вместо UDP-глюкозы в качестве промежуточного соединения образуется АТР-глюкоза)а
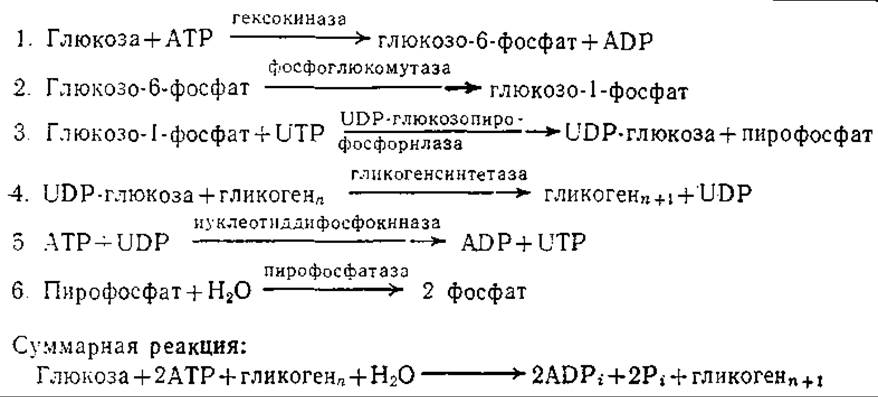
а Воспроизведено из работы: Lehninger A. L., Bioenergetics, 2d ed., р. 140, W. A. Benjamin, Inc., Раlо Alto, Ca., 1971.