Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Стехиометрия и энергетика метаболических превращений
Транспорт через клеточные мембраны
Активный транспорт
Как мы уже показывали на рис. 5.17, для активного транспорта характерно, во-первых, движение вещества против его химического (или электрохимического) градиента, т. е. из области с низкой концентрацией в область с высокой концентрацией, и, во-вторых, как показывает уравнение (5.41), необходимость затрат метаболической энергии. Например, если соединение, не несущее заряда, транспортируется из среды, где его концентрация равна 0,001М, в клетку, где его концентрация составляет 0,1 М, то
∆G°=1,98 кал/(моль∙град) (298 град) (ln 100) =
= 2,72 ккал/моль
Следовательно, для реализации этого процесса необходимо затратить по меньшей мере указанное количество энергии. Активный транспорт также осуществляется при участии специфических пермеаз.
Активный транспорт лежит в основе передачи нервных импульсов, но клетки используют этот механизм в гораздо более широких масштабах. Так, почти все клетки обладают системами активного транспорта (ионными насосами), позволяющими поддерживать необходимые концентрации ионов К+, Na+ и воды внутри клетки. Эти системы перемещают ионы Na+ из клетки, а ионы К+ в клетку, что позволяет компенсировать пассивную диффузию этих ионов. Указанные системы транспорта взаимосвязаны и сопряжены с гидролизом АТР (рис. 5.19). В красных кровяных тельцах роль натрий-калиевого насоса выполняет олигомерный белок, называемый Na+—К+-АТРазой. Как показано на рис. 5.19, этот белок располагается в толще плазматической мембраны и пронизывает ее насквозь; такие белки называют трансмембранными.
Вторая обычная группа систем активного транспорта обеспечивает поступление в клетку молекул питательных веществ, например глюкозы и аминокислот, со скоростями, значительно превышающими скорость их транспорта за счет пассивной диффузии. В клетках высших животных активный транспорт глюкозы совмещен с транспортом Na+ в клетку, а удаление Na+ из клетки осуществляется параллельно с гидролизом АТР под действием Na+—К+-АТРазы. Весь процесс схематично изображен на рис. 5.20. В бактериях для активного транспорта глюкозы используется другой процесс, называемый транслокацией групп. В этом процессе глюкоза высвобождается во внутриклеточное пространство в виде высокоэнергетического глюкозо-6-фосфата, с трудом проникающего через мембраны (рис. 5.21). Считается, что этот процесс является той стадией, скорость которой определяет скорость роста ряда клеток.
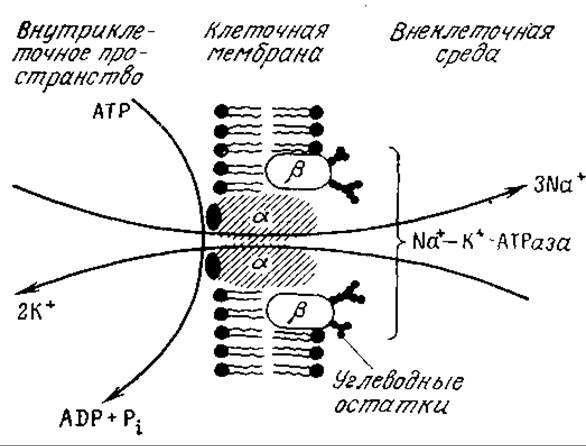
РИС. 5.19. Na+—К+-насос (Na+—К+-АТРаза) транспортирует Na+ из клетки и К+ в клетку; движущей силой этого процесса является гидролиз АТР.
Движущей силой многих систем транспорта в бактериях служит поток протонов через плазматическую мембрану. Основная роль потока протонов в энергетике бактерий описывается хемиосмотической теорией, разработанной Митчеллом в 1961 г. На рис. 5.22 изображена схема совместного транспорта протонов и лактозы в процессе дыхания бактерии Е. coli. Протоны и лактоза совместно транспортируются в клетку лактозопермеазой (продуктом транскрипции гена у оперона lac). Одновременно в дыхательной цепи протоны вместе с потоком электронов выводятся из клетки. В целом pH внутри клетки выше, чем в непосредственном ее окружении.
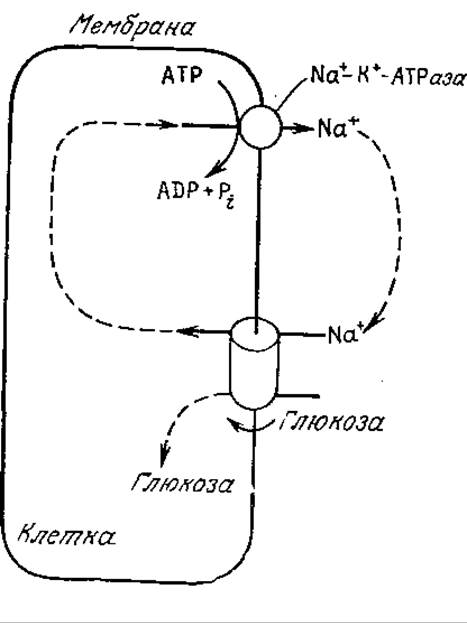
РИС. 5.20. Ионы Na+ и глюкоза транспортируются из среды в клетку одним и тем же пассивным переносчиком. Низкая внутриклеточная концентрация Na+ поддерживается за счет активного транспорта. (Воспроизведено с разрешения из работы: Lehninger A., Bioenergetics, 2d ed., р. 205, W. A. Benjamin, Inc., Palo Alto, CA, 1974.)
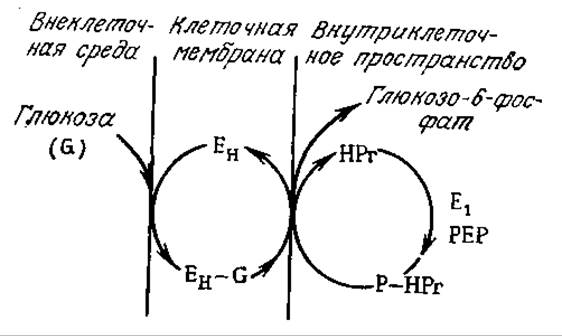
РИС. 5.21. Фосфорилирование глюкозы в клеточной мембране создает постоянную движущую силу для транспорта глюкозы через мембрану и удерживает глюкозу (в виде производного) внутри клетки. Здесь G — глюкоза; ЕH — мембранный фермент; E1— цитоплазматический фермент; НРr — цитоплазматический белок; РЕР — фосфоенолпируват. (Воспроизведено с разрешения из работы: Lehninger A. L., Biochemistry, 2d ed., р. 799, Worth Publishers, New York, 1975; есть перевод первого издания: Ленинджер А., Биохимия. — М.: Мир, 1976, с. 715.)
Эукариотические клетки должны обладать целым комплексом систем транспорта, поскольку концентрации многих веществ в органоидах поддерживаются на уровне, отличающемся от концентрации этих веществ в цитоплазме. Этот факт лишний раз говорит о том, что любая клетка, и особенно эукариотическая, представляет собой не однородную смесь множества компонентов, а высокоорганизованную, вплоть до молекулярного уровня, систему.
Разработано много математических моделей активного транспорта, но сейчас мы не будем их рассматривать. Более подробные сведения читатель может найти в приведенной в конце главы литературе и упражнениях.
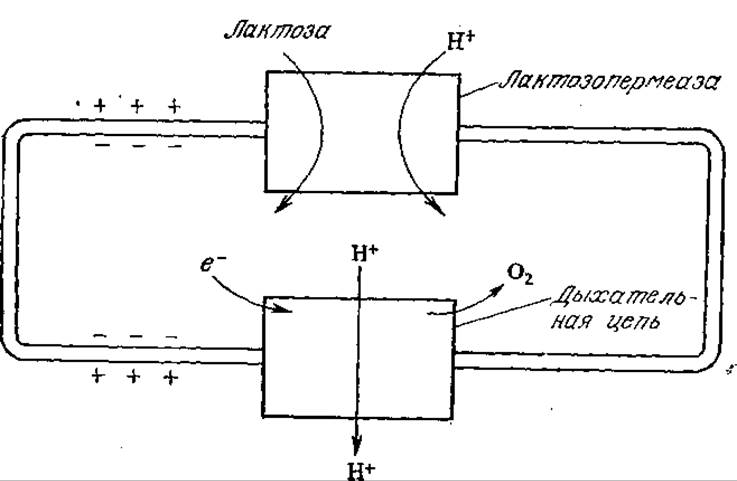
РИС. 5.22. Сочетание активного транспорта лактозы с транспортом протонов по градиенту электронного потока дыхательной цепи.
Прежде чем перейти к изучению потоков и регуляторных систем в последовательностях метаболических реакций, полезно несколько полнее и детальнее рассмотреть структуру и организацию клеточной мембраны. На рис. 5.23 схематично изображено строение клеточной оболочки бактерии; нетрудно видеть, что ее структура значительно сложнее описанной в гл. 2 упрощенной модели, включающей липидный бислой и белковую оболочку. На рисунке показано, что в толще мембраны находится множество белковых молекул; одни из них располагаются у наружной или внутренней поверхности мембраны, другие молекулы пронизывают всю толщу мембраны. Эти белки (пермеазы и белки-рецепторы) узнают специфические соединения в среде и контактируют с регуляторными системами клетки, обеспечивая ее ответную реакцию на изменения в среде. В случае бактерий белковые компоненты мембран позволяют клеткам быстро приспособиться к изменившимся условиям окружения. Поверхностные рецепторы высших эукариотических клеток участвуют в процессах клеточного узнавания, играющих большую роль как в организации тканей, так и в иммунной реакции клеток.
Мембраны в общем случае асимметричны; их наружные и внутренние поверхности содержат различные компоненты и выполняют различные функции. Липиды и разнообразные мембранные белки диффундируют в плоскости мембраны очень быстро; по некоторым оценкам скорость диффузии липидов в ряде мембран составляет 10-8 см2/с.

РИС. 5.23. Детали строения клеточной оболочки Е. coli. Здесь белые глобулярные объекты на поверхности и в толще клеточных мембран обозначают отдельные белки (НМ — наружная мембрана; ПГ — пептидогликан; ПП — периплазматическое пространство; ЦМ — цитоплазматическая мембрана). (Воспроизведено с разрешения из работы: Inouye М., Bacterial Outer Membranes, John Wiley, New York, 1979.)