Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Цикл лимонной кислоты
Цикл лимонной кислоты регулируется
Познакомимся теперь с тем, каким образом регулируется сам цикл лимонной кислоты (рис. 16-15). В большинстве случаев скорость функционирования метаболических циклов определяется их начальными этапами. Полагают, что так же обстоит дело и в случае цикла лимонной кислоты. Общая скорость его функционирования во многих тканях определяется первой реакцией:
Ацетил-СоА + Оксалоацетат → Цитрат + СоА.
Разумеется, скорость цитрат-синтазной реакции регулируется концентрацией ее субстратов, в частности концентрацией ацетил-СоА, а она в свою очередь зависит от активности пируватдегидрогеназного комплекса. Регулируется эта реакция также концентрацией второго субстрата - оксалоацетата; возможно даже, что этот фактор играет главную роль, поскольку концентрация оксалоацетата в митохондриях очень низка и зависит от метаболических условий. На активность цитрат-синтазы влияет также концентрация сукцинил-СоА, одного из более поздних промежуточных продуктов цикла. Как только концентрация сукцинил-СоА превышает нормальный стационарный уровень, цитрат-синтаза сразу же ингибируется, поскольку сукцинил-СоА понижает ее сродство к ацетил-СоА. Жирные кислоты, служащие предшественниками ацетил-СоА, тоже ингибируют цитрат-синтазу посредством аллостерических эффектов. В некоторых клетках роль ингибиторов цитрат-синтазы играют цитрат и NADH.
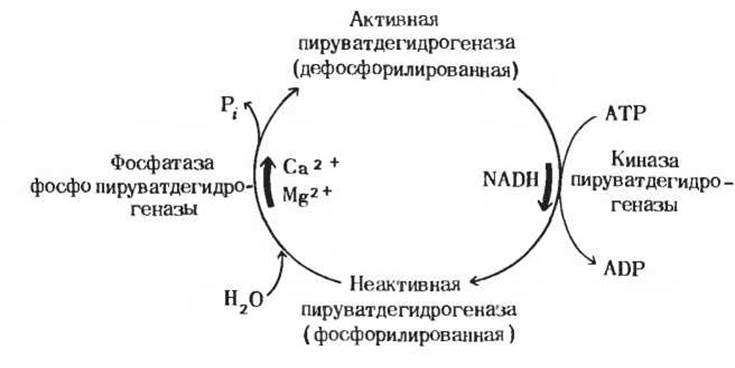
Рис. 16-14. Регуляция пируватдегидрогеназной реакции путем ковалентного взаимопревращения активной и неактивной форм фермента. Ионы Са2+ способствуют образованию активной формы фермента, стимулируя фосфатазу фосфопируватдегидрогеназы.
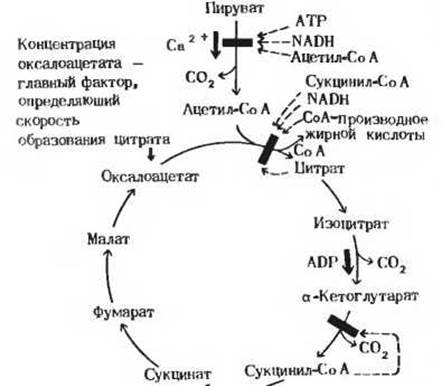
Рис. 16-15. Регуляция цикла лимонной кислоты при окислении пирувата в животных клетках. ATP, NADH, ацетил-СоА и Са2+ контролируют скорость образования ацетил-СоА из пирувата; скорость же функционирования цикла лимонной кислоты в целом регулируется концентрацией оксалоацетата, а также активностью цитрат-синтазы и изоцитратдегидрогеназы.
У большей части клеток окисление изоцитрата до а-кетоглутарата и СО2, которое может происходить под действием двух разных изоцитратдегидрогеназ, регулируется, по-видимому, путем аллостерической стимуляции NAD-зависимого фермента, вызываемой ADP. В то же время NADH и NADPH действуют как отрицательные модуляторы изоцитратдегидрогеназной активности. Ингибитором активности а-кетоглутаратдегидрогеназного комплекса служит продукт реакции сукцинил-СоА. Таким образом, в цикле лимонной кислоты регулируются по меньшей мере три стадии, и только в своих деталях эта регуляция у разных типов клеток несколько различается.
Скорость гликолиза в нормальных условиях согласована со скоростью функционирования цикла лимонной кислоты; в клетке до пирувата расщепляется ровно столько глюкозы, сколько необходимо для того, чтобы обеспечить цикл лимонной кислоты «топливом», т. е. ацетильными группами ацетил-СоА. Ни пируват, ни лактат, ни ацетил-СоА обычно не накапливаются в аэробных клетках в больших количествах; их концентрации поддерживаются на некоем постоянном уровне, соответствующем динамическому равновесию. Согласованность между скоростью гликолиза и скоростью функционирования цикла лимонной кислоты объясняется не только тем, что первый процесс ингибируется высокими концентрациями АТР и NADH, т.е. компонентами, общими для гликолитической и дыхательной стадий окисления глюкозы; определенную роль в этой согласованности играет также и концентрация цитрата. Продукт первой стадии цикла лимонной кислоты - цитрат — является аллостерическим ингибитором фосфофруктокиназы, катализирующей в процессе гликолиза реакцию фосфорилирования фруктозо-6-фосфата (разд. 15.13 и рис. 15.15).