Биохимия аминокислот - А. Майстер 1961
Промежуточный обмен аминокислот
Глицин, серин и саркозин
Взаимопревращение глицина и серина
В тканевом обмене млекопитающих и у микроорганизмов большое значение имеет процесс взаимопревращения глицина и серина. Шимин [165] впервые показал превращение серина в глицин при введении крысам и морским свинкам вместе с бензойной кислотой N15-серина, меченного С13 в карбоксильной группе; в выделенной из мочи животных гиппуровой кислоте содержание С13 в карбоксильной группе глицина было почти таким же, как и в исходном серине. Установлено также превращение глицина в серин [163]. Имеются многочисленные данные [166—170] об обратимом взаимопревращении глицина и серина, в котором участвует одноуглеродный остаток, представленный в следующей схеме в виде муравьиной кислоты:
![]()
Эту реакцию наблюдали в срезах печени и в целом организме крысы. В опытах на голубях было показано, что ß-углеродный атом серина может служить предшественником углерода уреидных групп мочевой кислоты [171]; известно, что эти углеродные атомы мочевой кислоты образуются из муравьиной кислоты. ß-Углеродный атом серина является также предшественником ß-углеродного атома этаноламина в липидной фракции печени [172]. Показано, что молочнокислые бактерии для синтеза серина из глицина и муравьиной кислоты нуждаются в пиридоксале [173]. Потребность в витамине В6 для этого процесса установлена также у цыплят. В опытах с препаратами печени В6-авитаминозных цыплят или цыплят, получавших предварительно дезоксипиридоксин, наблюдали снижение синтеза серина из глицина по сравнению с контрольными цыплятами [174, 175]. При добавлении к таким препаратам пиридоксальфосфата отмечали частичное восстановление активности.
1 Это превращение протекает через уропорфириноген III, который далее окисляется в уропорфирин III — общий предшественник порфириновых ядер гемина и хлорофилла. — Прим. ред.
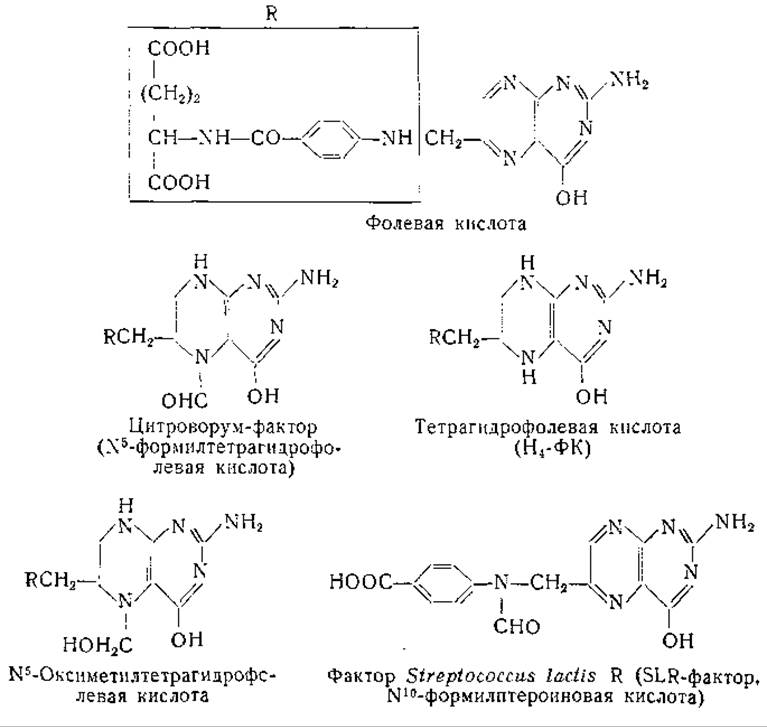
В ряде лабораторий [132, 174, 176—178] исследовали природу биологически активного одноуглеродного остатка, образующегося из муравьиной кислоты. Имеются данные о том, что при определенных условиях формальдегид используется для синтеза серина более активно, чем муравьиная кислота. Известно, что формальдегид является продуктом окисления саркозина и других N-метиламинокислот [179]; саркозин в свою очередь может служить источником одноуглеродных остатков для синтеза серина [178]. Между тем при добавлении немеченого формальдегида к полученной из печени крысы системе, образующей серин из меченой муравьиной кислоты, снижения радиоактивности в образующемся серине не наблюдали [176]. Эти, а также другие данные [180, 181, 1082] свидетельствуют о том. что активный одноуглеродный остаток не является ни муравьиной кислотой, ни формальдегидом. Это активное соединение, возможно, представляет собой промежуточный продукт обмена, общий для муравьиной кислоты и формальдегида [182]. Из различных исследований стало ясно, что в синтезе серина принимает участие фолевая кислота. Так, например, у крыс с недостаточностью фолевой кислоты оказалась пониженной скорость включения муравьиной кислоты в серин [183]. У различных животных при недостаточности фолевой кислоты затруднено также взаимопревращение глицина и серина [171, 184—187]. Данные, полученные при изучении микроорганизмов [173, 188, 189], также подтверждают участие производных фолевой кислоты в процессе взаимопревращения глицина и серина. Дальнейшим подтверждением служат данные, показавшие, что по крайней мере два производных фолевой кислоты («фактор Streptococcus lactis R» и «цитроворум-фактор») содержат формильные остатки и, таким образом, могут участвовать в переносе формильных групп [182, 190—200].
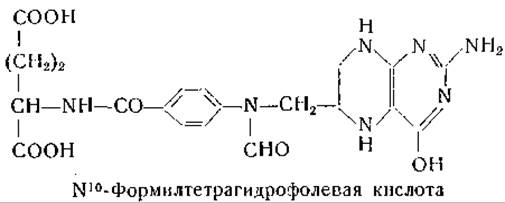
Это предположение было подтверждено в опытах с препаратом печени голубя, катализирующим процесс взаимопревращения серина и глицина. Обработка препарата печени голубя анионитом Дауэкс-1 в хлоридной форме с последующим диализом вела к инактивированию процесса включения С14-глицина в серин; при добавлении к таким препаратам тетрагидрофолевой кислоты отмечали восстановление активности [201]. Аналогичные результаты получены с ферментными препаратами из печени крысы [174, 202]. Тетрагидрофолевая кислота стимулировала также образование серина из глицина и формальдегида. Однако одна тетрагидрофолевая кислота не восстанавливает в инактивированных препаратах печени голубя способность к синтезу серина из глицина и муравьиной кислоты. В этой системе восстановление активности наблюдается при добавлении тетрагидрофолевой кислоты, аденозинтрифосфата, дифосфопиридиннуклеотида, глюкозо-6-фосфата и ионов магния [201, 202]. Из этих результатов можно заключить, что муравьиная кислота в присутствии АТФ переходит в производное цитроворум-фактора, которое при наличии в системе дифосфопиридиннуклеотида восстанавливается до оксиметилтетрагидрофолевой кислоты. Фолевая кислота проявляет некоторую активность в ферментной системе, использующей муравьиную кислоту для синтеза серина, но активность ее меньше, чем у тетрагидрофолевой кислоты. При синтезе серина из формальдегида и глицина дигидрофолевая кислота активирует ферментную систему в меньшей степени, чем тетрагидрофолевая кислота; если одновременно с дигидрофолевой кислотой добавляют АТФ, дифосфопиридиннуклеотид, глюкозо-6-фосфат и ионы магния, то процесс синтеза активируется в такой же мере, как и в пробах с тетрагидрофолевой кислотой. Эти данные показали, что в реакции превращения формальдегида в глицин и во включении глицина в серин участвует тетрагидрофолевая кислота. Для использования муравьиной кислоты в образовании серина из глицина необходимы дополнительные реакции.
Включение глицина в серин может происходить путем реакции обмена с участием тетрагидрофолевой кислоты в качестве кофермента, без увеличения общего количества серина. Роль витамина В6 в этой реакции состоит в активировании а-углеродного атома глицина (стр. 246), реагирующего с оксиметилтетрагидрофолевой кислотой; при расщеплении продукта реакции образуется серин. Саками [174] предложил для превращений одноуглеродных остатков следующую схему (см. также [203, 204]):
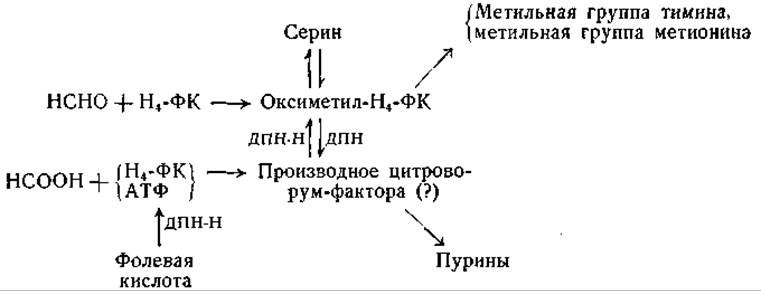
ß-Углеродный атом серина может переходить в метальные группы тимина и холина вместе со стоящими при нем водородными атомами [210—214] и может служить предшественником метальной группы метионина.
Енике [205] показал, что при переносе ß-углеродного атома серина в качестве первоначального продукта образуется N10-замещенное производное тетрагидрофолевой кислоты [206]. Этот автор доказал ферментативное образование N10-формил-тетрагидрофолевой кислоты и установил, что это соединение может служить донатором одноуглеродного остатка при синтезе серина, пуринов и гистидина. Согласно этой точке зрения, N15-формилтетрагидрофолевая кислота рассматривается как продукт побочной реакции. Гринберг и сотрудники [207] показали, что N10-формилтетрагидрофолевая кислота участвует непосредственно в синтезе пуринов путем переноса формильного остатка на 5-амино-4-имидазолкарбоксамид-5-фосфорибозид. Результаты исследований, проведенные с системой из печени голубя, могут быть выражены следующими уравнениями [1083]:
![]()
Для реакции (1) необходимы фосфопиридоксаль и ионы марганца; реакция (2) протекает медленнее с ДПН, чем с ТПН.
Опубликованы данные, согласно которым превращение серина в глицин в экстрактах одного из видов Clostridium происходит в присутствии дифосфопиридиннуклеотида, ионов марганца, пиридоксальфосфата, ортофосфата и нового фактора, обозначенного как кофермент С. Этот фактор отличается от упомянутых выше производных фолевой кислоты. Из С. cylindrosporum были выделены 5 групп птеридиновых соединений, обладающих активностью коферменте С; оказалось, что некоторые из них содержат глутаминовую кислоту, глицин, серин и аланин [208, 209]. Имеются указания на то, что в обмене одноуглеродных соединений может участвовать витамин Е [215]. Так, например, при введении кроликам с недостаточностью витамина Е C14-муравьиной кислоты последняя включалась в нуклеиновые кислоты и белки значительно более активно, чем у контрольных животных; если вводили 1-С14-глицин, то у животных с недостаточностью витамина Е включение изотопа было понижено.
Найдено, что в растении табака формальдегид и ß-углеродный атом серина используются как предшественники N-метильной группы никотина, причем в этом процессе формальдегид более эффективен [216—218]. Метальная группа метионина может служить предшественником метоксильных групп лигнина, являясь в этом превращении значительно более эффективным донатором, чем муравьиная кислота [219]. а-Углеродный атом гликолевой кислоты может служить источником как N-метильной группы никотина, так и метоксильных групп лигнина [1084].
Серин участвует также в реакции синтеза триптофана из индола и серина; эта реакция рассматривается в разделе, посвященном обмену триптофана.