Биохимия - Химические реакции в живой клетке Том 1 - Д. Мецлер 1980
Мембраны и клеточные оболочки
Метаболизм и функции мембран
Проведение нервных импульсов
Способность мембран поддерживать разность концентраций ионов между цитоплазмой и внеклеточной средой обусловливает их возбудимость, что имеет важное практическое значение. Разделение зарядов создает как бы «аккумулятор» легко доступной энергии, которая может быть использована для проведения электрических сигналов через клеточную мембрану. Эта способность клеток проводить сигналы проявляется в наиболее яркой форме в нервных волокнах, однако она свойственна не только нервным клеткам и обнаруживается даже у простейших, например у Paramecium [70, 70а], а также, вероятно, у бактерий.
В настоящее время еще мало известно о химической природе проведения нервных импульсов, однако электрические характеристики этого процесса детально изучены и описаны. Если ввести в клетку через мембрану микроэлектрод, то можно измерить разность потенциалов между внешней средой и содержимым клетки. Эта разность потенциалов, получившая название потенциала покоя, достигает в нервных клетках 90 мВ. Своим происхождением она обязана, по-видимому, различиям в концентрациях ионов. Исходя из величины ∆G для разбавления раствора иона [уравнение (3-25)] и соотношения между ∆G и электродным потенциалом [уравнение (3-63)], можно легко получить уравнение Нернста:
![]()
Согласно этому уравнению, которое применимо к системе, содержащей ионы только одного типа, при 10-кратной разнице концентраций однозарядного иона (n=1) по разные стороны от мембраны будет возникать потенциал, равный 59 мВ. Поскольку мембраны относительно мало проницаемы для ионов натрия, принято считать, что мембранный потенциал образуется главным образом за счет разницы концентраций ионов калия. Наряду с уравнением (5-2) получено также и более полное уравнение, учитывающее как концентрации ионов К+, Na+ и Сl-, так и их способность проникать через мембрану [69, 71, 72].
Если на каком-нибудь участке мембраны проницаемость для ионов натрия увеличивается, то эти ионы устремляются внутрь клетки, нейтрализуя ее отрицательный заряд. Клеточная мембрана при этом деполяризуется. При деполяризации по поверхности мембраны распространяется затухающий электрический сигнал, аналогично тому как это имеет место при прохождении тока по коаксиальному кабелю Считают, что включение неровного импульса часто связано с локальным увеличением проницаемости мембраны для ионов натрия. В этом процессе могут играть определенную роль также и другие ионы, в частности Са2+. Пассивное распространение электрических сигналов, обусловленное локальной деполяризацией мембраны, происходит, однако, только в случае очень коротких нервных клеток; на длинные расстояния этим способом сигнал распространяться не может. В большинстве аксонов нервных клеток используется более эффективный способ проведения импульса, основанный на развитии потенциала действия. Потенциал действия — это импульс, проходящий вдоль аксона и специфически изменяющий за доли секунды (в нервах млекопитающих приблизительно за 0,5 мс) мембранный потенциал (рис. 5-6). Исходный отрицательный потенциал ~50—70 мВ быстро падает до нуля, затем достигает положительного значения 40—50 мВ, после чего снова устанавливается потенциал покоя. Поразительная особенность потенциала действия состоит в том, что он распространяется вдоль аксонов со скоростью 1—100 м/с без снижения интенсивности.
Для изучения химической природы потенциала действия в 50-х годах А. Ходжкин и А. Хаксли разработали метод фиксации потенциала. С помощью этого остроумного метода можно измерять трансмембранный ток, поддерживая мембранный потенциал на требуемом уровне с помощью системы, работающей по принципу обратной связи [69, 71, 73]. Использование фиксации потенциала позволило измерять зависимость проводимости мембраны от мембранного потенциала и от времени. Оказалось, что сразу же после того, как с помощью фиксации потенциала мембранный потенциал устанавливается на пониженном уровне, проницаемость мембраны для ионов натрия резко возрастает. Увеличение проницаемости автоматически приводит к деполяризации прилежащей области мембраны и соответственно к образованию самораопространяющейся волны, движущейся вдоль аксона. Химическая природа процессов, изменяющих проницаемость мембраны, остается неясной. С помощью фиксации потенциала было установлено, что через доли миллисекунды проницаемость мембраны возрастает также и для ионов калия. При этом проницаемость для ионов натрия снова уменьшается, и через некоторое время вновь восстанавливается обычный мембранный потенциал1. Последовательность событий в этом процессе можно представить следующим образом сначала открываются натриевые «каналы» (это не то же самое, что поры в Na+-насосе), а затем — калиевые каналы, после чего каналы закрываются в той же последовательности Результаты этих исследований позволили Ходжкину и Хаксли вывести уравнения, позволяющие количественно оценивать потенциалы действия и предсказывать наблюдаемые на опыте скорости проведения и ряд других характеристик нервных импульсов.
Отличительным свойством нервных волокон, обладающих способностью очень быстро проводить импульсы, является наличие миелиновой оболочки. Миелиновые мембраны надежно изолируют аксон от окружающей среды на всем протяжении, кроме специальных участков, перехватов Ранвье, которые располагаются по ходу аксона на расстоянии 1—2 мм друг от друга За счет этого нервный импульс обретает способность как бы перескакивать с одного участка на другой. Такое сальтаторное (скачками) проведение происходит со значительно большей скоростью (до 100 м/с) по сравнению со скоростью проведения в немиелинизированных аксонах.
Что же известно о каналах, проводящих ионы Na+ и К+ при возбуждении нерва? С помощью тетродотоксина иглобрюхих (рис. 16-7) было убедительно показано, что для этих двух ионов существуют раздельные каналы. Этот вывод был сделан на основании того, что токсическое действие тетродотоксина сопровождалось блокированием натриевых каналов, тогда как проведение ионов К+ при этом не нарушалось. В то же время калиевые каналы блокировались при помощи различных четвертичных аммониевых солей. Высокое значение константы связывания тетродотоксина (приблизительно 3∙108 М-1) позволило определить число натриевых каналов путем титрования. Хотя данные, полученные разными автора ми, различались, тем не менее даже максимальные значения свидетельствуют об относительно небольшом числе натриевых каналов — 40—75 на 1 мкм2 поверхности мембраны [74] (для сравнения напомним, что в мембране на той же площади расположено 2∙106 фосфолипидных молекул). Число каналов, проводящих ионы натрия, по-видимому, в 10 раз меньше числа «насосных» каналов, т. е. каналов, используемых (Na+ + К+) -зависимой АТРазой [75]. Число калиевых каналов не известно
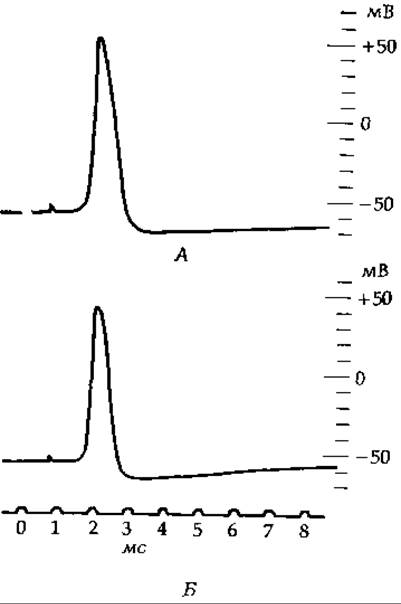
РИС 5-6 А Потенциал действия, записанный с помощью электрода, введенного в аксон, который был предварительно освобожден от цитоплазмы, а затем заполнен раствором сульфата калия (18°С). Б. Потенциал действия интактного аксона, записанный с тем же усилением и той же временной шкалой (18°С). На шкале напряжения указана разность потенциалов между внутренним электродом и таким же электродом, погруженным во внешний раствор (без поправки на контактный потенциал) (Hodgkin А, Conduction of Nervous Impulses, 1964; с любезного разрешения Charles С. Thomas, Publisher, Springfield, Illinois)
1 Однако в период абсолютной рефрактерности (длящейся приблизительно 0,5 мс) проведение нервного импульса невозможно.
Поскольку число каналов, проводящих ионы, мало, скорость прохождения натрия через открытые каналы должна быть чрезвычайно высокой; по оценкам, она равна ∼108 ионов в 1 с. Отсюда следует, что эти каналы не могут работать с помощью ионофорных переносчиков, а должны представлять собой просто «поры», открывающиеся и закрывающиеся в соответствии с изменениями мембранного потенциала. Было высказано предположение, что, прежде чем пройти через каналы, образованные, судя по всему, встроенными в мембрану белковыми молекулами, ионы калия и натрия избавляются от сольватной оболочки.
Как открываются «ворота» ионных каналов? Характер роста скорости проводимости при изменении мембранного потенциала явно указывает на кооперативный характер этого процесса [74, 76]. Вряд ли можно что-либо добавить в настоящее время к тому, что оказано в этом разделе о проведении нервных импульсов. Выяснение химических основ ионной проводимости в возбудимых мембранах является одной из самых актуальных задач биохимии.