Биохимия - Химические реакции в живой клетке Том 1 - Д. Мецлер 1980
Молекулы, из которых мы состоим
Сахара и полисахариды
Полисахариды
Полимеры сахаров присутствуют во всех клетках и выполняют множество функций. Так, целлюлоза придает прочность зеленым растениям, хитин обусловливает прочность скелета членистоногих. Гиалуроновые кислоты и другие мукополисахариды образуют защитную прослойку между животными клетками, а пектины и родственные полисахариды играют аналогичную роль в растениях. Клеточные поверхности обычно покрыты слоем полисахаридов самой разной структуры. Различия в структуре полисахаридов, составляющих этот наружный слой, весьма важны, поскольку обусловливают иммунологическую индивидуальность организмов. Крахмал, гликоген и другие запасные полисахариды представляют собой легко мобилизуемые пищевые ресурсы клеток [35 а].
Один из компонентов растительного крахмала, амилоза, — это линейный полимер, состоящий из многих a-D-глюкопиранозных звеньев, которые соединены друг с другом посредством 1,4-связи. В гранулах крахмала всегда содержится и второй компонент, амилопектин2. И амилопектин, и гликоген (животный крахмал) состоят из сильно разветвленных молекул. Ветви присоединены к а-1,4-цепям посредством а-1,6-связей (рис. 2-14 и 2-15). Некоторые бактерии, например Leuconostoc mesenteroides, синтезируют линейную (1—>6)-поли-D-глюкозу, или декстран; ряд видов присоединяет ветви посредством а-1,4- или а-1,3-связей. Декстраны, синтезируемые бактериями, обитающими на поверхностях зубов, являются компонентами налета на зубах. Декстраны производятся в промышленном масштабе; после химической обработки, «наделяющей» их множеством поперечных связей, образуются гели (коммерческое название их «сефадекс»), широко используемые в биохимических лабораториях для разделения смесей разных молекул (разд. 3.1.г).
Целлюлоза — это, пожалуй, самый распространенный на земле углевод (растения производят ее в количестве ~1014 кг в год); он представляет собой полиглюкозу, образованную с помощью ß-1,4-связей. Хитин, другой весьма распространенный полисахарид, содержится в клеточных стенках грибов и в наружном скелете членистоногих. Это линейный ß-1,4-полимер N-ацетилглюкозамина, структурно сходный с целлюлозой. Пектины представляют собой ß-1,4-пoлигaлaктypoнoвыe кислоты, в которых ряд карбоксильных групп метилирован.
Клеточные стенки дрожжей содержат полимеры маннозы (маннаны); в них от главной а-1,6-цепи отходят короткие ветви (из 1—3 маннозных звена), соединенные а-1,2- и а-1,3-связями. Кроме целлюлозы, все растения содержат ксиланы, которые состоят преимущественно из ß-1,4-ксилопиранозных цепей. Отметим, что ксилоза, представляющая собой пятиуглеродный сахар, в этом полимере принимает форму шестичленного пиранозного кольца. С другой стороны, фруктоза, шестиуглеродный сахар, в инулине, запасном полисахариде иерусалимского артишока, присутствует в форме пятичленных фуранозных колец. Эти различия обусловлены различиями в путях биосинтеза. Пиранозные кольца термодинамически более устойчивы, чем фуранозные. Последние образуются в инулине и сахарозе, поскольку их биосинтез проходит через 6-фосфатный эфир фруктозы.
2 Некоторые виды крахмала, в частности крахмал «восковой» кукурузы, содержат один лишь амилопектин и не содержат амилозы.
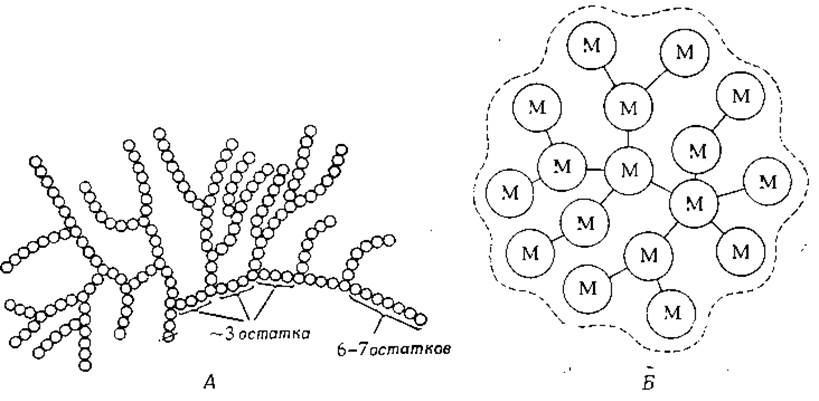
РИС. 2-15. Схематическое изображение молекулы гликогена. А. Структура участка молекулы гликогена по Мейеру (Meyer К- Н., Adv. Enzymol., З, 109—136, 1943). Кружки обозначают остатки глюкозы, соединенные а-1,4-связями или (в точках ветвления) а-1,6-связями. Перечеркнутый кружок обозначает редуцирующую группу. [French D.r in: Symposium on Foods: Carbohydrates and Their Roles (H. W. Schulz ed.), pp. 26— 54. Avi Publ., Westport, Connecticut, 1969.] Б. Двумерная схема молекулы гликогена с мол. весом 760 000, содержащей ~4700 остатков глюкозы (штриховая линия — внешние контуры молекулы). При обработке ß-амилазой (гл. 7, разд. В.6) получается «остаточный декстрин» с мол. весом ~410 000, который на рисунке представлен кружками и соединяющими их линиями. Буквами М обозначены сильно ветвящиеся участки, которые соединены друг с другом участками с малым числом точек ветвления. Под. действием а-амилазы (гл. 7, разд. В.6) М-участки частично разрушаются с образованием остаточных декстринов, содержащих в среднем по 34 остатка глюкозы. (Brammer G. A., Rougvie М. A., French D., Carbohyd. Res., 24, 343—354, 1972.)
Ряд полисахаридов, включая различные виды крахмала, содержит один редуцирующий конец, что обусловливает возможность раскрытия кольца и образования свободной альдегидной группы, обладающей восстановительными свойствами. В других случаях редуцирующий коней, «заблокирован». Один из видов такой блокировки (реализуемой, как предполагается, в инулине) состоит в том, что конец цепи имеет структуру сахарозы. В других полисахаридах возможно присутствие концевой трегалозы. Встречаются циклические полисахариды, сахарные цепи которых замкнуты в кольцо и вообще не имеют свободного редуцирующего конца. Эти полисахариды, как правило, присоединены к молекулам белка или липида.
Во многих полисахаридах повторяющимися единицами являются мономеры разного типа. В некоторых из таких гетерополисахаридов: имеет место простое чередование двух сахаров [36, 37]. Примерами служат гиалуроновая кислота, хондроитинсульфаты и дерматансульфат — важные компоненты основного вещества, присутствующего в межклеточном пространстве соединительной ткани и выполняющего роль цементирующего материала. Гиалуроновая кислота — это полимер, в котором чередуются глюкуроновая кислота и N-ацетилглюкозамин; структура этого полимера изображена на рис. 2-16. Хондроитин-сульфаты и дерматансульфат имеют аналогичное строение, но в первом случае в состав полисахарида входит N-ацетилгалактозамин, а во втором — а-L-идуроновая кислота; кроме того, в положениях, указанных на рис. 2-16, находятся сульфоэфирные группы.
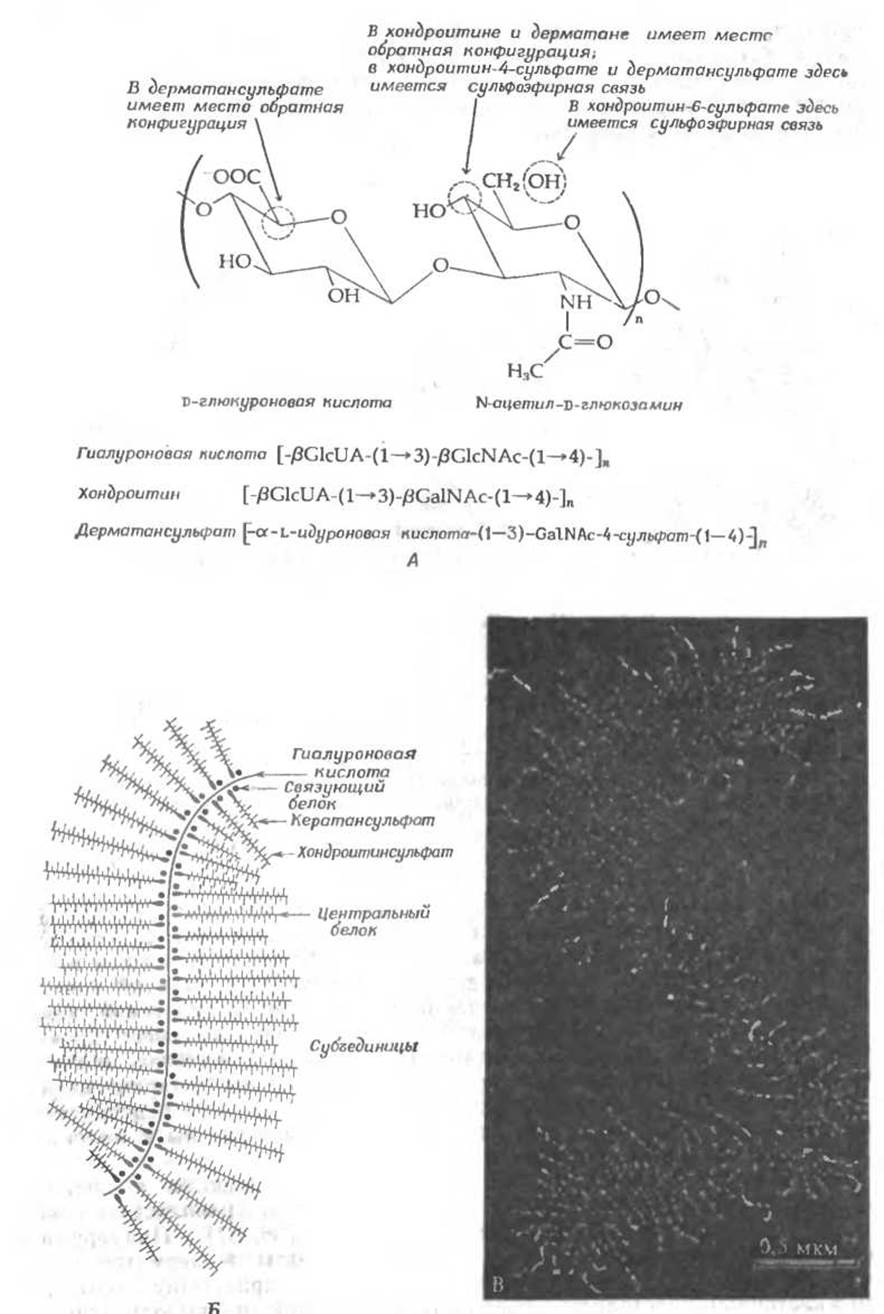
РИС. 2-16. А. Повторяющиеся дисахаридные звенья гиалуроновой кислоты и родственных мукополисахаридов. Б. Предполагаемая схема строения протеогликановых агрегатов в хрящевых тканях, на которой указано расположение молекулы гиалуроновой кислоты, а также хондроитинсульфата и кератансульфата. Последние ковалентно связаны с белком, образующим центральную часть субъединицы протеогликана; единый протеогликановый агрегат образуется путем присоединения этих субъединиц к молекуле гиалуроновой кислоты. Рисунок выполнен с примерным соблюдением пропорций, за исключением того, что длины цепей хондроитинсульфата изображены почти наполовину укороченными. [Rosenberg L., in: Dynamics of Connective Tissue Macromolecules (P. M. C. Burleigh and A. R. Poole, eds.), p. 107, North-Holland, Amsterdam, 1975.] Такие мукополисахарндные цепи могут электростатически взаимодействовать с находящимся в хрящах коллагеном. В. Электронная микрофотография (негативный контраст) протеогликанового агрегата из бычьего суставного хряща (расположенного на несущих поверхностях суставов). Волокнистый остов состоит из гиалуроновой кислоты (см. рис. Б), от остова отходят протеогликановые субъединицы (индивидуальные полисахаридные цепи не видны). [386]. (С любезного разрешения L. Rosenberg.)
Гепарин представляет собой мукополисахарид с антикоагуляционными свойствами, секретируемый в кровь тучными клетками, присутствующими в легких, печени и других тканях. Этот линейный полисахарид, по всей вероятности, содержит дисахаридные звенья следующего типа: [—уроновая кислота-(1—>4)-GlcN-2,6-дисульфат- (1—>4)—]n.
Обе аминогруппы и 6-гидроксилы остатков глюкозамина несут сульфатные группы. В некоторых звеньях встречается D-глюкуроновая кислота, присоединенная посредством а-1,4-связи, однако наиболее часто в роли первого звена дисахарида выступает (L-идуроновая кислота)-2-сульфат [38].
Кератаны кожи в качестве повторяющегося звена содержат [—ßGal-(1—>4)-ßGlcNAc-6-сульфат-(1—>3)—]n или точно такую же структуру, но содержащую галактозаминсульфат. Из-за присутствия карбоксилатных и сульфатных групп все эти полимеры несут большое число отрицательных зарядов.