Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Коферменты - особые природные специализированные реагенты
Флавиновые коферменты
Комплексы металлов с флавинами и металлофлавопротеиды
Наличие ионов металлов в некоторых флавопротеидах [125а] свидетельствует о непосредственном присоединении ионов металлов к флавинам. Хотя окисленные флавины связывают ионы большинства металлов нелегко, они образуют окрашенные в красный цвет комплексы с Ag+ и Сu+ с диссоциацией протона от N-3 [126, 127]:
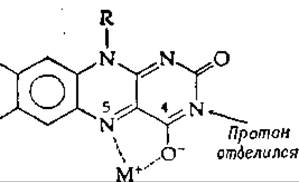
Флавиновые семихиноны (радикалы) образуют более прочные красные комплексы со многими металлами [128]. И в этом случае полагают, что ион металла взаимодействует с N-5 и атомом кислорода при С-4. Если тот же центр занят ионом металла, имеющим более чем один уровень окисления, то передача электронов между флавином и субстратом может протекать при посредстве атома металла.
Необычный флавопротеид ксантиноксидаза катализирует две реакции (8-56 и 8-57).
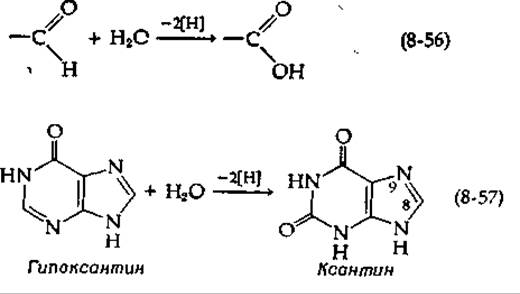
Близкородственный фермент альдегидоксидаза осуществляет только реакцию (8-56). Ксантиноксидаза может также окислять ксантин дальше, путем повторения окислительного процесса того же типа в положениях 8 и 9 [см. реакцию (8-57)] с образованием мочевой кислоты. Возможно, наиболее интересное свойство ксантиноксидазы заключается в том, что на каждую молекулу связанного FAD она содержит прочно связанный атом молибдена (дополнение 14-А) наряду с четырьмя атомами железа. (Молекула является димером с мол. весом ~ 275 000, содержащим две молекулы FAD, два атома Мо и восемь атомов Fe.)
Фактически первым указанием на роль молибдена в метаболизме послужило открытие того обстоятельства, что недостаточность молибдена в рационе животных приводит к уменьшению активности ксантиноксидазы в печени (фермент содержится и в молоке).
Реакции (8-56) и (8-57) записаны так, как если бы ксантиноксидаза являлась дегидрогеназой: сначала вода присоединяется к связи С — О — или к C=N в том месте, где происходит окисление, после чего аддукт подвергается дегидрированию. Такой механизм не объясняет потребности в Мо и Fe.
Обнаружение характерных ЭПР-сигналов молибдена в процессе действия фермента указывает на то, что Мо действительно участвует в катализе [129, 129а]. Первым этапом ферментативной реакции может быть восстановление Mo(VI) в Mo(V). Гамильтон предположил наличие механизма [117], показанного в уравнении (8-58).
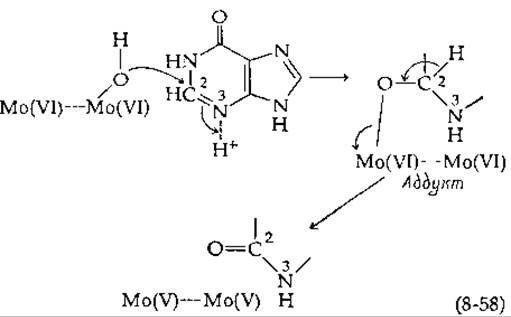
Аддукт, возникающий первоначально в результате присоединения координированной с металлом гидроксильной группы, может далее окисляться переносом электронов через кислород на два атома молибдена. Каждый из них мог бы получать один электрон с образованием двух атомов Mo(V). Электроны могут далее переноситься на флавины, на связанные с белками атомы железа и на О2 по миниатюрной цепи переноса электронов. Ксантиноксидаза принадлежит к группе флавиновых ферментов, содержащих прочно связанное железо, не координированное с ядром гема. Она является представителем железо-серных белков, которые обсуждаются дальше, в гл. 10 (разд. В). Ксантиноксидаза и альдегидоксидаза содержат также надсульфидную группу (—S—S-), которая имеет существенное значение для проявления каталитической активности.
К флавинсодержащим железо-серным белкам принадлежит NADH-дегидрогеназа (реакция д, табл. 8-4) и сукцинатдегидрогеназа [реакция (8-49)]. Оба этих митохондриальных фермента переносят электроны, вероятно, при помощи связанных атомов железа на систему цитохромов цепи переноса электронов (гл. 10) [130а].
Некоторые металлофлавопротеиды содержат группы гема (гл. 10, разд. Б). Примером может служить L-лактатдегидрогеназа дрожжей — фермент, который также называют цитохромом b2. Этот белок является тетрамером с мол. весом ~ 235 000; каждая его субъединица состоит «з двух различных полипептидных цепей, одной молекулы рибофлавин- фосфата и одного гема [131]. По всей вероятности, флавин и гем прикреплены к различным полипептидным цепям и фермент действует как типичная дегидрогеназа, причем электроны передаются от восстановленного флавина через атом железа на связанный гем. Внешним акцептором является, по-видимому, цитохром с.