Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Рост, дифференцировка и химическая коммуникация клеток
Нейрохимия
Обмен веществ в нейронах и химия процесса мышления
Нейроны характеризуются необыкновенно высоким уровнем обмена веществ, значительная часть которого направлена на обеспечение работы натриевого насоса в мембранах и поддержание состояния возбуждения. Химические основы передачи нервного импульса по аксону уже обсуждались в гл. 5, разд. Б, 3. Последовательное раскрытие сначала натриевых и затем калиевых каналов можно считать твердо установленным. Менее ясным остается вопрос, сопряжено ли изменение ионной проницаемости, необходимое для распространения потенциала действия, с какими-либо особыми ферментативными процессами. Нахманзон указывает, что ацетилхолинэстераза присутствует в высокой концентрации на всем протяжении мембраны нейрона, а не только в синапсах [38, 39]. Он предполагает, что увеличение проницаемости к ионам натрия обусловлено кооперативным связыванием нескольких молекул ацетилхолина с мембранными рецепторами, которые либо сами составляют натриевые каналы, либо регулируют степень их открытия. При этом ацетилхолин высвобождается из участков накопления, расположенных на мембране, в результате деполяризации. Собственно, последовательность событий должна быть такова, что изменение электрического поля в мембране индуцирует изменение конформации белков, а это уже приводит к высвобождению ацетилхолина. Под действием ацетилхолинэстеразы последний быстро распадается, и проницаемость мембраны для ионов натрия возвращается к исходному уровню. В целом приведенное описание отличается от описанной ранее схемы синаптической передачи только в одном отношении: в нейронах ацетилхолин накапливается в связанной с белками форме, тогда как в синапсах — в специальных пузырьках. Существует мнение, что работа калиевых каналов регулируется ионами кальция. Чувствительный к изменению электрического поля Са-связывающий белок высвобождает Са2+, который в свою очередь активирует каналы для К+; последнее происходит с некоторым запозданием относительно времени открытия натриевых каналов, что обусловлено различием в константах скоростей этих двух процессов [123]. Закрытие калиевых каналов обеспечивается энергией гидролиза АТР. Имеются и другие предположения о механизмах нервной проводимости [124]. Некоторые из них исходят из того, что нервная проводимость целиком обеспечивается работой натриевого насоса.
Вернемся к такой специфической особенности нейронов, как высокая скорость обмена веществ. Ядро и большая часть рибосом расположены в теле нервной клетки. Однако многие белки необходимы в высокой концентрации в аксоне и синаптических окончаниях. К таким белкам относятся ферменты синтеза и распада нейромедиаторов, а также мембранные белки. При перерезке аксона отделенное синаптическое окончание очень скоро атрофируется; это наблюдение еще много лет назад позволило заключить, что из тела клетки на периферию поступают какие-то необходимые вещества. Экспериментально установлено, что действительно многие соединения перемещаются от тела клетки вниз по аксону со скоростью 1—10 мм/день. Больший интерес, однако, представляет недавно открытый быстрый транспорт в аксоне. Оказалось, что белки и другие вещества движутся со скоростью до 0,4 м/день. Винбластин (дополнение 4-А) и батрахотоксин (рис. 12-18) специфически блокируют этот вид транспорта. Высказано, предположение, что белок, обладающий АТРазной активностью и химически сходный с головками миозина, плюс тонкие нити и микротрубочки составляют функциональную систему, которая образует своего рода канал, или, скорее, миниатюрные рельсы, для транспорта вещества вдоль микрофиламентов [125, 126]. Перенос веществ может идти и в противоположном направлении, т. е. от синаптических окончаний к телу клетки. Это так называемый ретроградный транспорт по аксону, благодаря которому, вероятно, изменяются свойства нейрона в ответ на электрическую активность синаптических окончаний [127].
Имеются данные, свидетельствующие о том, что в клетках головного мозга транскрибируется значительно большая часть генома, чем в других клетках [128, 129]. Так, ~20% ДНК мозга человека гибридизируется с мРНК, синтезированной в клетках мозга. В клетках других органов и тканей транскрибируется примерно вдвое меньше ДНК. У человека количество транскрибируемой ДНК выше, чем у мыши [128]. В связи с этим следует упомянуть о том удивительном факте, что в мозге человека и мыши нет общих по электрофоретической подвижности форм ферментов [129]. Значение этих факторов пока неясно.
Какие химические процессы лежат в основе мышления и создают поток сознания в мозге человека? Поступление импульсов в мозг оказывает большое влияние на сигналы, идущие на периферию по моторным нейронам. Известно также, что мозг обладает собственными эндогенными электрическими ритмами, которые не зависят от импульсов, поступающих по сенсорным нейронам. У примитивных беспозвоночных источником таких ритмов служат особые нейроны — водители ритма (пейсмейкеры). Эти нейроны спонтанно возбуждаются с постоянными интервалами. По-видимому, в их клеточных мембранах происходят последовательные циклические изменения ионной проницаемости, достаточные для возникновения потенциала действия. Примеры работы трех типов нейронов — водителей ритма у моллюсков [130] приведены на рис. 16-12. Вполне вероятно, что аналогичный феномен лежит в основе работы мозга человека. Вероятно,- сознательная мысль возникает при сочетании ритмов от эндогенных водителей ритма с импульсацией, поступающей от сенсорных нейронов. Возвращаясь к примитивным организмам, любопытно сравнить спонтанный ритм нейронов — водителей ритма с периодическим выбросом сАМР клетками Dictyostellium (гл. 6. разд. 5). Может быть, эти два феномена по существу имеют много общего.
Чрезвычайно любопытны вопросы, касающиеся химической основы памяти. Если процесс мышления осуществляется путем прохождения каких-то систем электрических волн через сеть нейронов в коре мозга, то где и в какой форме накапливаются следы этого процесса, или энграммы памяти? Как показали эксперименты, существуют кратковременная память с относительно малой способностью к накоплению и долговременная память. Накопленная информация может переходить из кратковременной формы запоминания в более длительную. Считается, что кратковременная форма памяти представлена реверберирующими контурами, возникающими в коре больших полушарий и быстро исчезающими. Кратковременная память может полностью исчезнуть, например, после удара по голове. Долговременная память, напротив, сохраняется в течение столь длительного времени, что ее можно связывать с появлением каких-то устойчивых изменений в химизме нейронов, или даже в физических связях между ними.
Учитывая, что передача нервных импульсов осуществляется по принципу «все или ничего», разумно допустить, что именно в синапсах происходят те изменения, которые ведут к запоминанию. При исследовании отдельных синапсов были обнаружены явления облегчения синаптической передачи и привыкания (торможения). Феномен облегчения состоит в том, что второй импульс, как правило, передается через синапс более эффективно, чем первый; привыкание же состоит в снижении ответа на повторяющиеся стимулы. Память в сущности может оказаться комбинацией этих двух феноменов в специфических синапсах. Вспомним, что возбуждение мускариновых рецепторов в холинэргических синапсах стимулирует высвобождение в клетках cGMP, тогда как возбуждение под влиянием катехоламинов стимулирует образование сАМР. Циклические нуклеотиды в свою очередь регулируют активность множества внутриклеточных ферментов. Это ведет к изменению не только свойств синаптических мембран, но и транскрипция генов и многих других процессов. В результате зависящего от сАМР фосфорилирования увеличивается активность РНК-полимеразы II, а также орнитиндекарбоксилазы, участвующей в образовании полиаминов (гл. 14, разд. В, 4). Таким образом, прохождение импульса через синапс должно оказывать продолжительное влияние на свойства этого синапса.
Целый ряд экспериментальных данных подтверждает существование химической основы памяти. Например, введение животным небольших доз стрихнина облегчает обучение [131]. Другие вещества, например пуромицин (рис. 15-18), оказывают противоположное действие [129, 132]. Процесс обучения у животных связан с увеличением синтеза в нейронах мРНК и белков. Существенно важно, что синтез полипептидов и нуклеиновых кислот протекает в основном в теле нервной клетки, а не в окончаниях аксонов или в дендритах. Тело нервных клеток покрыто обычно синаптическими пуговками, и вполне вероятно, что именно стимуляция поверхности мембран тела клетки индуцирует синтез макромолекул.
В отличие от приведенной гипотезы, придающей основное значение в механизме обучения явлениям облегчения нервной передачи и привыкания, другая гипотеза рассматривает в качестве химической основы обучения молекулярный код. Действительно, из мозга крыс, приученных избегать темноты, был выделен пептид, состоящий из 15 аминокислотных остатков, связанный с указанным поведенческим навыком: при введении пептида в мозг необученных крыс они также начинали избегать темноты [133]. Это только один пример из множества сообщений о существовании в мозге специфических переносчиков приобретенных навыков поведения. Все же учитывая сложность структуры мозга и имеющиеся сведения о механизмах функционирования нейронов, гипотезу молекулярного кода довольно трудно принять. В то же время ее нельзя полностью отбросить. В самом деле, существование пептидных гормонов и либеринов, синтезируемых в нейронах (разд А, 1), заставляет внимательно отнестись к предположению о связи долговременной памяти с синтезом специфических аминокислотных последовательностей в определенных нейронах.
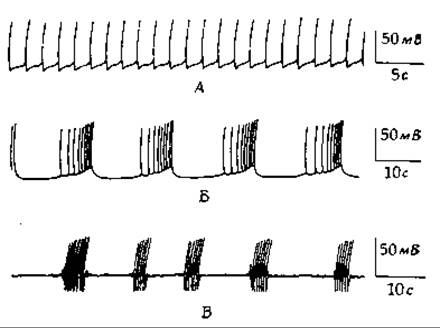
РИС. 16-12. Электрическая активность изолированных нейронов моллюска Aplysia. А. Ритмический водитель ритма. Б. «Разрядный» водитель ритма. В. Осциллирующий водитель ритма [130].
Обучение связано с различными частями мозга. Структура мозжечка хорошо изучена, и эту часть мозга можно рассматривать как орган, где протекают процессы запоминания и обучения. Имеются теории относительно химической природы этих процессов в мозжечке [134].