Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Рост, дифференцировка и химическая коммуникация клеток
Дифференцировка тканей и биология развития
Физиологическая модуляция и различные программы развития
Процесс развития животного из оплодотворенного яйца — одно из наиболее замечательных биологических явлений. Из первых, очень сходных между собой эмбриональных клеток в ходе всего нескольких клеточных делений возникают дифференцированные органы и ткани, такие, как печень, мозг, почки, кожа и эритроциты. Дифференцированные клетки характеризуются, как правило, высокоспециализированными биохимическими свойствами. Так, эритроциты содержат гемоглобин, тогда как в мышечных клетках в больших количествах образуются миозин и актин. В эндокринных клетках поджелудочной железы синтезируются инсулин и глюкагон, а в экзокринных — пищеварительные ферменты, которые секретируются в пищеварительный тракт. В целом считается, что в клетках специализированных тканей одновременно транскрибируется не более 10% общего количества генов (исключение составляет ткань мозга; см. разд. Б, 8). Методом химического анализа четко установлено, что специализированные клетки содержат нормальное количество ДНК, т. е. полный набор генов, но 90% этого количества не функционирует.
Хотя в химических основах механизма дифференцировки клеток еще много неясного, все же известно, что в этом процессе исключительно важную роль играют химические сигналы, поступающие из внешней среды и от прилегающих клеток. Эти сигналы запускают внутреннюю, генетически детерминированную программу развития, определяющую путь дифференцировки отдельных клеток. С какой точностью выполняется программа развития, можно показать на примере коловраток и кольчатых червей (рис. 1-10), отдельные виды которых характеризуются почти непогрешимым постоянством числа клеток. Так, у нематоды Oxyuris equi имеются точно 251 нервная клетка, одна экстреторная клетка, 18 клеток средней кишки и 64 мышечные клетки [140].
Прежде чем обсуждать вопрос о дифференцировке сложных многоклеточных организмов, полезно рассмотреть более примитивные формы — одноклеточные и колониальные. В благоприятных условиях клетки бактерий и эукариот одинаковым образом вступают в фазу «роста и деления» (рис. 15-25), которая составляет основу экспоненциального роста [уравнение (6-60)]. Однако изменение внешних условий быстро меняет характер жизнедеятельности клеток. Так, недостаточность питательного субстрата не только уменьшает скорость роста, но и влияет на транскрипцию генов. У Е. coli это происходит в результате увеличения концентрации в клетке сАМР. Наличие дополнительного источника энергии, например лактозы, также может индуцировать специфические изменения в транскрипции генов (гл. 15, разд. Б, 1). Можно привести множество других примеров, свидетельствующих о влиянии физиологических факторов на генетический аппарат (так называемое явление «физиологической модуляции»).
Еще более удивителен тот факт, что под влиянием факторов внешней среды в клетке может быть запущена иная программа развития, связанная с активацией дополнительных генов и существенной перестройкой всей клетки. Примером может служить образование спор у некоторых бактерий (гл. 1, разд. А, 8), происходящее при неблагоприятных для вегетативного роста условиях внешней среды.
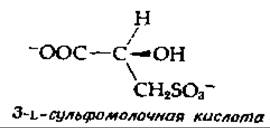
Целый ряд химических сдвигов происходит при споруляции [141— 143]. Вначале полностью прекращается синтез рРНК, далее начинается транскрипция новых классов мРНК и образуется несколько новых белков. Обращает на себя внимание образование в больших количествах дипиколиновой кислоты (рис. 14-7), что требует появления по крайней мере одного нового фермента. Кроме того, в процессе образования спор бактерии потребляют много Са2+, значительные количества Мn2+ и ионов других металлов. У многих бактерий образуется также 3-L-сульфомолочная кислота.
В спорах Bacillus subtilis указанные вещества содержатся в следующих количествах (в расчете на сухой вес): дипиколиновая кислота — 10%, сульфомасляная кислота — 3—6%, Са2+—3%, Мn2+—0,3%. Споры бактерий удивительно устойчивы к нагреванию и не теряют жизнеспособности при длительном выдерживании в кипящей воде. Распространено мнение, что именно дипиколиновая кислота и другие ионы придают белкам устойчивость к денатурации. Устойчивость к нагреванию может быть связана также с высокой степенью обезвоженности спор [144]. В условиях, благоприятных для роста, споры прорастают и бактериальные клетки снова переходят на программу, обеспечивающую рост и деление.
Переключения на более сложные программы развития наблюдаются у колониальных форм бактерий, например у миксобактерий, но химические сигналы, вызывающие переключение, еще неизвестны [145]. Выявлено, однако, что у относящихся к эукариотам миксомицетов типа Dictyostelium (гл. 6, разд. Е, 5), имеющих такую же программу развития, сигналом субстратного голодания служит выброс сАМР1). Повышение концентрации сАМР воспринимается другими клетками, у которых в ответ так изменяются процессы биосинтеза, что происходят дифференцировка и образование плодовых тел [135, 136, 146]. Отдельные клетки начинают вырабатывать целлюлозу, а также мукополисахариды; образуется трегалоза, которая накапливается в спорах. Синтезу этих продуктов предшествует образование новых ферментов.
1) Нельзя исключить, что чувствительность к сАМР возникает как вторичный ответ на выделение какого-то макромолекулярного фактора, стимулирующего дифференцировку [144а].
У водной плесени Blastocladiella, относящейся к фикомицетам, образование тонкостенного или же толстостенного устойчивого к нагреванию спорангия определяется анионом НСО-3, т. е. более простым химическим фактором, чем сАМР. С другой стороны, у примитивных эукариот роль химических сигналов могут играть макромолекулярные соединения, по всей вероятности белки. Так, некоторые штаммы Dictyoslelium переходят на другую генетическую программу, а именно начинают образовывать макроцисты путем конъюгации двух различных типов клеток под влиянием диффундирующего индуцирующего фактора с мол. весом — 12 000, выделяемого, по-видимому, клетками одной из линий [147].
Благодаря относительно простому строению некоторые ткани растений служат удобным объектом изучения процесса дифференцировки. Слой камбия в стебле (рис. 1-12) постоянно дифференцируется с образованием флоемы из наружно расположенных клеток и ксилемы из клеток, расположенных со стороны сердцевины стебля. В то же время часть камбиальных клеток сохраняется недифференцированными. Фактически при каждом клеточном делении одна дочерняя клетка подвергается дифференцировке, тогда как другая остается малодифференцированной камбиальной клеткой. Такой способ постоянной дифференцировки стволовых клеток, сохраняющих постоянные свойства, широко распространен как у растений, так и у животных. По-видимому, направление дифференцировки камбиальных клеток зависит от химической природы сигналов, которые идут от клеток, прилегающих к камбию с наружной или внутренней стороны. Известно, что к числу факторов, индуцирующих дифференцировку, относятся сахароза, ауксин и цитокинины.
Обычно в основе вегетативного размножения растений лежит способность эмбриональной ткани меристемы (гл. 1, разд. Д.4) дифференцироваться в корни и побеги. С другой стороны, при культивировании изолированных клеток флоемы или других дифференцированных тканей, как правило, формируется так называемый каллус, т. е. масса претерпевших дифференцировку клеток, напоминающих эмбриональные. При создании благоприятных условий, в частности при культивировании в среде, содержащей кокосовое молоко, а также при соблюдении соответствующего соотношения концентраций ауксина и цитокинина удавалось индуцировать реверсию, т. е. превращение клеток флоемы корня моркови в эмбриональные клетки, из которых затем развивалось целое растение [136]. Этот опыт имеет принципиальное значение, так как определенно доказывает, что дифференцированные клетки флоемы моркови содержат полный набор генов, необходимых для развития растения. Вместе с тем существенно и то, что с большинством растений такого рода опыт воспроизвести довольно трудно и процесс дедифференцировки далеко не всегда происходит автоматически. Все же это происходит в достаточном числе случаев, чтобы установить факт тотипотентности ядра дифференцированных клеток.
Данные многочисленных экспериментов указывают, что дифференцировка частично возникает как реакция клетки на химические сигналы, поступающие от соседних клеток или из внешней среды.