Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Рост, дифференцировка и химическая коммуникация клеток
Дифференцировка тканей и биология развития
Гормональная регуляция роста
Отдельные органы образуются, как правило, из клеточных закладок эктодермы и энтодермы. Многочисленные данные свидетельствуют, что такие закладки образуются под влиянием химических веществ, выделяемых клетками прилежащего зародышевого слоя. Так, формирование из эктодермы невральной пластинки, из которой в будущем разовьется головной мозг, и хорды индуцируется мезодермальными клетками, лежащими непосредственно под областью невральной пластинки. Аналогичным образом в ходе взаимодействия клеток мезо- и эктодермы возникают молочные железы, тогда как образование поджелудочной железы, печени и легких определяется взаимодействием группы клеток энтодермы и мезодермы. Участие специфических химических агентов в таких взаимодействиях доказывается тем, что эффект индукции можно наблюдать в условиях, исключающих непосредственный контакт между клетками, например если разделить клетки тонкими (20 мкм) миллипоровыми фильтрами [150].
Имеющиеся данные позволяют считать, что большое число химических соединений различных типов участвует в межклеточных взаимодействиях, функционируя в качестве гормонов местного действия, или аутокоидов. Эти вещества индуцируют рост или дифференцировку, служат факторами хемотаксиса, некоторые из них несомненно тормозят рост. Приведем несколько фактов.
Поразительный пример химической индукции процесса развития можно наблюдать, используя сенсорные и симпатические нейроны эмбриона. Интенсивный рост аксонов эмбриональных нейронов (рис. 16-14) индуцируется особым фактором роста нервов [151—153], который представляет собой небольшой белок, состоящий из 118 аминокислот и содержащий три дисульфидные связи. По величине и общей структуре этот белок напоминает проинсулин; в функциональном отношении его следует рассматривать как истинный гормон. Он синтезируется во многих тканях, но больше всего в подчелюстных (слюнных) железах мышей-самцов. Эти железы, подобно поджелудочной железе, содержат как эндокринную, так и экзокринную ткань. Гормон роста нервов секретируется также фибробластами; кроме того, он обнаружен в злокачественных клетках саркомы. Предполагается, что в самой нервной системе гормон продуцируется клетками глии. Механизм его действия заключается, по-видимому, в том, что он изменяет свойства мембран нервных клеток, способствуя быстрому отрастанию длинных тонких аксонов. Имеется сообщение о существовании другого белка с большим молекулярным весом, действующего на клетки мозга эмбриона примерно таким же образом [154].
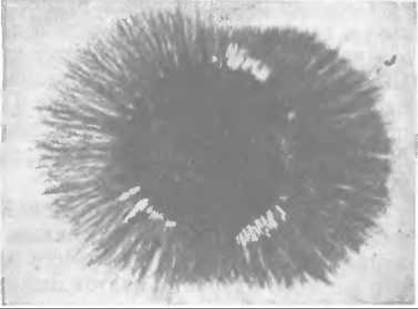
РИС. 16-14. Влияние 1 нг фактора роста нервов на образование аксонов сенсорный ганглием куриного эмбрина (Frazier W. A., Oblendorf С. Е., Boyd L. Е., Aloe L., Johnson E. M., Ferrendelli J. A., Bradshaw R. A., PNAS, 70, 2448—2452 (1973).
При изучении структуры мозга, а также формирования в ходе эмбрионального развития соединений между нейронами бросается в глаза одно удивительное обстоятельство [155]. Дело в том, что весь мозг пронизан множеством связей еще до того, как он начал функционировать, и в ходе образования такой «проводки» аксоны прорастают из одной части мозга в другую, протягиваясь иногда на очень большие расстояния. Но еще более удивительно, что соединения между нейронами формируются строго постоянным образом. Например, изображение, возникшее на сетчатке глаза, без помех передается в специфическую область мозга, где оно воспринимается. Нейроны, осуществляющие связь между сетчаткой и мозгом, растут так, будто их снабдили точным адресом того участка мозга, с которым они должны соединяться [155]. В настоящее время нет решительно никаких данных, какая химически закодированная информация лежит в основе этого явления.
Подобно фактору роста нервов, существует эпидермальный фактор роста, также синтезирующийся в подчелюстной железе мышей-самцов; это полипептид, который состоит из 53 аминокислотных остатков [156, 157]. Выделена свойственная человеку форма этого фактора [157], идентичная, видимо, урогастрону [158]. Последний обладает свойством предотвращать развитие язвы желудка и находится в относительно больших количествах в моче беременных женщин (у которых обычно не развиваются язвы).
Подобно тому как отростки нейронов растут и соединяются со специфическими участками-мишенями, так и целые клетки нередко направленно перемещаются в процессе эмбрионального развития. Полагают, что в основе миграции клеток лежит хемотаксис (разд. Б. 7). Значительная часть работ по изучению миграции клеток выполнена на гидре (рис. 1-10), примитивном животном, содержащем клетки только 10 типов. Один из этих типов представлен эмбриональным резервом клеток мезодермы. Это клетки стволовой линии, образующие, помимо прочих клеток, нематоциты (стрекательные клетки), которые, сформировавшись, продвигаются вверх по телу гидры и располагаются в конце концов в щупальцах [159, 160].
Ранее уже упоминалась роль хемотаксиса в функционировании защитной системы человеческого организма (дополнение 5-Ж). Этому несколько противоречит постулат, что каждая клетка продуцирует тканеспецифичный гормон местного действия, называемый келоном, который ингибирует митотическую активность других клеток той же ткани. Полагают, что эти гормоны играют важную роль в регуляции деления клеток и предотвращении злокачественного роста. Выделено несколько келонов, оказавшихся белками или пептидами различного молекулярного веса [161, 162].
В целом гормоны местного действия оказывают различное действие на близлежащие клетки, в частности усиливают или тормозят рост, влияют на миграцию, а также на дифференцировку клеток. Во многих случаях гормоны местного действия — это специфические пептиды или белки, но в этом качестве могут выступать также более простые соединения: небольшие пептиды, гистамин, серотонин и даже бикарбонат- ион [163]. Эти вещества, передвигаясь от одной клетки к другой, должны проникать через клеточные мембраны. Обычно они переносятся жидкостью, окружающей клетки, но иногда и более прямым путем — через щелевые соединения (гл. 1, разд. Д, 3, а). В этих соединениях имеются специальные каналы, по которым происходит транспорт между цитоплазмой соседствующих клеток низкомолекулярных веществ, по-видимому белков [164, 165, 165а]. Еще одна форма строго направленного пути межклеточных коммуникаций — это синапсы.