Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Рост, дифференцировка и химическая коммуникация клеток
Дифференцировка тканей и биология развития
Программы развития у многоклеточных организмов
Как мы уже видели, клетки постоянно получают химические сигналы как непосредственно от прилегающих клеток, так и через омывающие жидкости; в ответ на это они высвобождают определенные соединения либо так или иначе меняют свойства своей поверхности. Возникает, однако, вопрос, могут ли в ходе такого межклеточного взаимодействия сформироваться 200 типов специализированных клеток, свойственных организму млекопитающих. Тот факт, что даже бактериальные клетки могут переключаться с одной программы развития на другую, делает такое предположение вероятным. У низкоорганизованных животных на определенном этапе развития яйцеклетки синтез ДНК выключается и в клетке начинают накапливаться большие количества РНК, которая используется в дальнейшем эмбриональном развитии. На ранних стадиях эмбрионального развития основную организующую роль играют такие факторы, как полярность яйцеклетки и градиент концентрации всех ее компонентов. Следовательно, ядра яйцеклеток отвечают на внешние стимулы таким образом, что обеспечивают исходную полярность эмбриона. На самых ранних стадиях развития процесс дифференцировки легко обратим. В дальнейшем же превращение дифференцированной клетки в клетку эмбрионального типа становится трудным или даже невозможным. Опыты Гёрдона (разд. В, 2 данной главы) показывают, что ядро дифференцированной клетки обычно (если не всегда) содержит весь генетический материал. Этому факту нисколько не противоречат многочисленные экспериментальные данные, свидетельствующие о том, что на ранних стадиях развития клетки, расположенные в разных частях зародыша, следуют различной внутренней генетической программе так, словно направление дифференцировки у них предопределено. В некоторых случаях создается впечатление, будто заводятся некие «часы развития», которые полностью определяют дальнейший ход дифференцировки.
Это представление подтверждается существованием стволовых клеток, сохраняющих некоторые черты эмбриональных клеток; при каждом делении стволовой клетки образуется новая стволовая клетка плюс дифференцированная клетка. Последнее явление трудно объяснить только как реакцию на химические сигналы из окружающей среды. Согласно некоторым наблюдениям, клетки животных обладают ограниченным потенциалом деления [176, 177]. Например, нормальные диплоидные фибробласты человеческого эмбриона при выращивании в культуре делятся примерно 50±10 раз, после чего погибают независимо от условий культивирования. Фибробласты, полученные от людей Старшего возраста, погибают после меньшего числа клеточных делений. Аналогичным образом быстрее погибают в культуре клетки животных с меньшей продолжительностью жизни, например клетки мышей (после 14—28 делений) [177]. Из этих наблюдений можно сделать вывод о существовании внутренней программы, предопределяющей время гибели клеток. Имеются, однако, наблюдения, противоречащие этому выводу [178], так что в целом концепция о наличии верхней границы числа делений дифференцированных клеток остается недоказанной.
Третья линия данных в пользу существования внутренних программ развития вытекает из тщательно проведенных эмбриологических исследований. В частности, показано, что у куриного эмбриона зачаток конечности (длина которого равна сумме диаметров 20 клеток) содержит, клетки, совершенно автономно дифференцирующиеся в дальнейшем в отдельные элементы органа. Если эту зону развития с одного зачатка конечности перенести на второй симметричный зачаток, то там разовьется конечность, содержание костных и хрящевых элементов в которой будет в два раза больше нормального [179].
Современные теории развития принимают существование определенных генетических программ и рассматривают весь процесс развития как результат сочетания реакций клетки на воздействие гормонов и индукторов с влиянием внутренней генетической программы [179]. В настоящее время можно высказать только первые догадки о природе внутренних программ. Все же были предложены очень разумные схемы, согласно которым часы развития считают число клеточных делений и в соответствующий момент выключают одни гены и включают другие [180]. Были высказаны конкретные предположения относительно химизма таких часов. Так, указывалось, что вопреки представлению о высокой стабильности ДНК это соединение легко мутирует под влиянием химических факторов. Можно допустить существование особых ферментов, направленно модифицирующих ДНК в определенных участках. В самом деле, известно, что в ДНК содержится определенное количество дополнительных метальных групп, которыми, например, могут быть маркированы отдельные участки (гл. 2, разд. Г, 8). Другая возможность — это дезаминирование содержащих аминогруппу оснований в определенных участках, например в палиндромных последовательностях.
Вполне возможно, что происходит следующая цепь событий. Под действием соответствующего фермента аденин в паре АТ может быть дезаминирован в инозин. В результате после деления клетки одна из, дочерних клеток получит неизмененную молекулу ДНК, тогда как во второй клетке вместо пары оснований АТ окажется пара UC. При следующей репликации возникнет пара GC. Таким образом, в части дочерних клеток в специфическом участке ДНК происходит замена АТ на GC. Такая простая замена, возникающая под действием особого фермента, образовавшегося на определенной стадии развития, может изменить в некоторых клетках выражение отдельных генов. Вполне вероятно, что другой фермент способен вызвать обращение указанного эффекта, т. е. превратить модифицированную пару оснований в исходную форму. Например, дезаминирование цитозина и последующая репликация ДНК приведут к образованию пары AU, которая после второй репликации ДНК превратится в исходную пару АТ. Если специфические палиндромные участки доступны и многократно повторяются, то можно себе представить, что действие модифицирующего фермента последовательно распространяется по всей длине хромосом в обоих направлениях. Именно таким образом может возникнуть эффект включения специфических генов после определенного числа клеточных делений (подробности см. в работе Холидея и Пуга [1801).
На рис. 16-16 показано, как гипотетический фермент E1 может модифицировать участок ДНК путем метилирования основания в одной из двух палиндромных последовательностей. Этот фермент, которому приписывается довольно необычная специфичность, должен также метилировать второй участок в комплементарной цепи, но вне палиндромной области. После репликации одна молекула ДНК остается неизмененной, тогда как вторая окажется субстратом фермента Е2. Под действием последнего произойдет метилирование второй половины палиндрома и всех ДНК-потомков. В результате совместного действия ферментов E1 и Е2 модифицированные клетки будут все больше и больше отличаться от немодифицированных, т. е. будет идти процесс дифференциации, подобный тому, какой имеет место в стволовых клетках, подвергающихся дифференцировке. Несколько иная схема (но также предполагающая наличие фермента довольно необычной специфичности) объясняет механизм внутреннего счета числа клеточных делений аналогично постулату о транзициях АТ — GC[180].
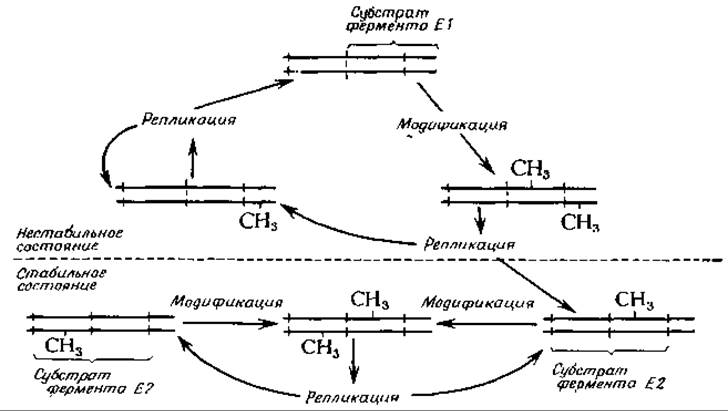
РИС. 16-16. Отделение метилированной ДНК от неметилированного предшественника. Первый модифицирующий фермент E1 метилирует половину палиндромной последовательности и прилегающую последовательность в комплементарной цепи. В ходе репликации образуется субстрат для второго фермента Е2, который метилирует вторую половину палиндрома и все последующие поколения молекул ДНК. При наличии E1 и Е2 из немодифнцированных или частично модифицированных клеток постоянно образуются стабильные модифицированные клетки. В случае инактивации или исчезновения E1 образуются стабильные модифицированные и иемодифицнрованные клетки (Holliday R., Pugh J. Е., Science, 187, 227, 1975).
Как же тогда объяснить тотипотентность ядер дифференцированных клеток? Имеются многочисленные данные о том, что в цитоплазме яйцеклетки содержатся факторы, выключающие транскрипцию специализированных генов. Создается впечатление, что какой-то механизм переводит стрелки часов развития, заставляя клетки дифференцироваться. Вполне вероятно, что до тех пор, пока не происходит заметной потери ДНК из генома, модифицированная ДНК ферментативно превращается в исходную немодифицированную форму. Если рассматривать метилированную ДНК, то весьма существенно, что в случае отсутствия в цитоплазме яйцеклетки ферментов E1 и Е2 (рис. 16-16) дальнейшего метилирования в ходе дробления не произойдет. На стадии гаструлы, когда, видимо, начинают включаться часы развития, в ДНК большинства клеток метилированный основания должен отсутствовать.