Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Свет в биологии
Поглощение света веществом
Круговой дихроизм и дисперсия оптического вращения
Круговой дихроизм образца обусловлен различием между молярными коэффициентами экстинкции для право- и левополяризованного света [уравнение (13-7]; он наблюдается только для хиральных молекул:
![]()
Значение ∆ε может быть непосредственно измерено с помощью дихрографа. Спектр кругового дихроизма (КД) часто напоминает спектр поглощения данного образца, поскольку положение пиков для обоих случаев совпадает. Однако КД бывает как положительным, так и отрицательным, причем он может быть положительным для одного перехода и отрицательным для другого (рис. 13-8). Все более широкое применение находят графики, представляющие зависимость Де от длины волны или волнового числа. Часто в литературе используется молярная эллиптичность:
![]()
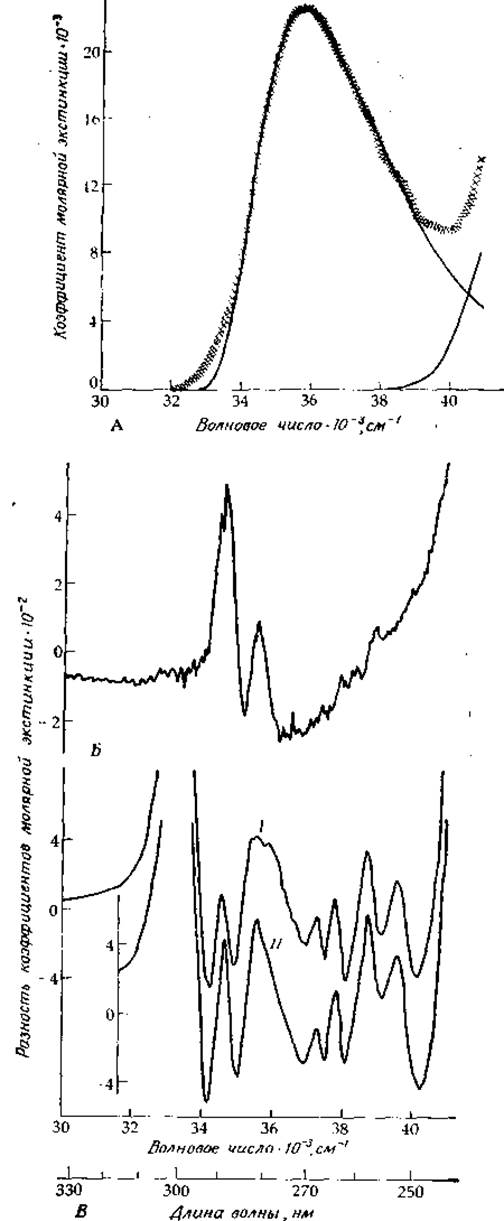
РИС. 13-13. А. Спектр поглощения каталитической субъединицы аспартат—карбамоил-трансферазы (крестики) с вписанными в него логарифмически-нормальными кривым» (сплошные линии). Б. Разность между спектром поглощения фермента в присутствии 0,09 М сукцината и 4,3 мМ карбамоилфосфата и спектром А (ср. с разностным спектром поглощения интактной аспартат—карбамоилтрансферазы, приведенным в работе [39]). В. Кривая I: «тонкая структура», выявляющаяся после вычитания спектра А из сглаженной кривой — результата суммирования двух логарифмически-нормальиых кривых. Кривая ІІ: тот же спектр для фермента+сукцинат+карбамоилфосфат. Фермент был предоставлен Дж. Нагелем н Г. Шахманом, а спектр сиял И.-И. Янг.
Вращательная сила полосы в спектре КД определяется выражением
![]()
Интегрирование производится по всей полосе поглощения, отвечающей данному переходу.
Круговой дихроизм тесно связан с дисперсией оптического вращения — изменением оптического вращения с длиной волны. Оптическое вращение обусловлено различием в показателях преломления (nL— nR) для лево- и правополяризованного света. Вращение а измеряется в градусах или радианах. Обычно все данные представляют в виде удельного вращения, т. е. вращения по отношению к вращению гипотетического раствора, содержащего 1 г ∙ мл-1 вещества в кювете длиной 1 дм. Зная вращение, концентрацию с' (в г∙мл-1) и длину кюветы I' (в дм), можно рассчитать удельное вращение по формуле
![]()
Молекулярное вращение задается выражением (13-11):
![]()
Здесь М — молекулярный вес, а с и I измеряются соответственно в моль∙л-1 и в см. Чтобы учесть небольшую поправку на поляризуемость поля, действующего на молекулы, вводят множитель 3/(n2 + 2). Вращение в радианах на 1 см оптического пути можно непосредственно выразить через длину волны и показатели преломления nL и nR:
![]()
В отличие от кругового дихроизма дисперсия оптического вращения (ДОВ) распространяется в спектральную область, далекую от полос поглощения образца. По мере приближения к полосе поглощения оптическое вращение возрастает либо в положительном, либо в отрицательном направлении. Затем в пределах самой полосы поглощения оно резко падает до нуля и далее принимает противоположный знак (коттон-эффект). Хотя наличие оптического вращения в той области, где вещество не поглощает, является определенным преимуществом метода ДОВ, интерпретировать спектры ДОВ несколько сложнее. В принципе данные, полученные с помощью указанных методов, взаимозависимы, и в обоих случаях мы получаем химическую информацию одного и того же типа. Поскольку регистрация спектров КД и ДОВ не составляет труда, а также благодаря чувствительности этих спектров к конформационным изменениям и к изменениям состава среды, оба метода широко используются в биохимии.
В качестве примера рассмотрим спектр КД медьсодержащего белка (рис. 13-8). КД в области d—d-полос спектра поглощения меди отчасти обусловлен асимметрией окружения иона меди в структуре белка. Такова же причина и нередко наблюдаемого кругового дихроизма ароматических аминокислот белков. Для тирозина знак КД может быть как положительным, так и отрицательным, но при этом он остается постоянным для всей полосы поглощения. Вследствие этого полосы КД по форме сходны с полосами поглощения [19, 46]. Фенилаланин ведет себя сложнее. Колебательные полосы, следующие за 0—0-полосой с интервалом 930 см-1, имеют одинаковый знак, а их интенсивности соотносятся с интенсивностью 0—0-полосы так же, как и в спектре поглощения. Однако колебания, волновые числа которых равны волновому числу 0—0-полосы плюс 180 и 520 см-1, иногда приводят к появлению полос КД противоположного знака, с различным соотношением между интенсивностями [22, 46]. Таким образом, спектр КД может оказаться более сложным и труднее поддающимся интерпретации, чем спектр поглощения того же хромофора.
Нередко связывание симметричного хромофора с молекулой белка или нуклеиновой кислоты приводит к появлению КД у этого хромофора. Например, связанный ферментом пиридоксальфосфат или пиридоксаминфосфат (рис. 8-8) имеет положительную полосу в спектре КД, тогда как полоса хиноноидного промежуточного соединения с Vmax = 20 400 cм-1 (490 нм) характеризуется отрицательным КД. Это замечательное явление наводит на мысль, что спектр КД хромофора содержит определенную информацию о его окружении, однако никакой простой интерпретации здесь пока найти не удается.
Легче интерпретировать дихроизм n—п*-переходов карбонильных соединений. В данном случае имеется набор правил, известных как правила октанта, которые позволяют предсказывать знак и величину КД простых соединений [47]. Разработан также теоретический подход к анализу КД-спектров и спектров поглощения белков в высокоэнергетической УФ-области. В пределах регулярной ß-структуры, а-спирали и кристаллических областей электронные переходы соседствующих друг с другом амидных групп могут быть связаны, в результате чего имеет место делокализация возбуждения. Такая делокализация (экситон) приводит к расщеплению (давыдовскому расщеплению) на два перехода с различающимися энергиями и направлением поляризации [7, 44]. Так, полоса поглощения амидной группы с Vmax = 52 600 см-1 в случае а-спирали расщепляется на две компоненты с Vmax = 48 500 и 52 600 см-1. Кроме того, низкоэнергетические п—п*- и n—п*-переходы весьма близки по энергии, что может приводить к формированию состояния, представляющего смесь двух указанных состояний с появлением вращательной силы в п—п*-полосе, знак которой противоположен знаку вращательной силы в п—п*-полосе (см. работу [44]). И знак, и интенсивность КД-полос зависят от конформации соединения, что позволяет четко различать а-спирали, ß-структуры и статистический клубок. В водных растворах измерения проводят при длинах волн, простирающихся вплоть до вакуумного ультрафиолета, т. е. до волновых чисел ~60 000 см-1 [48].
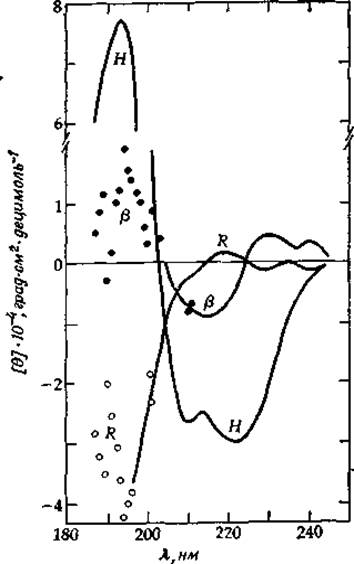
РИС. 13-14. Спектры для спирали (Н), ß-структуры и неупорядоченного клубка (R), рассчитанные по спектрам КД пяти белков. В той области, где не удается провести гладкую кривую, оставлены точки. [Chen Y.-H. et al., Biochemistry, 13, 3353 (1974).]
При всех достижениях теоретического характера по предсказанию формы КД-спектров более ценным часто оказывается эмпирическое сопоставление спектров разных соединений. Например, на рис. 13-14 приведены КД-спектры спиралей, ß-структур и неупорядоченных пептидных цепей, рассчитанные из измеренных спектров в сочетании с анализом реальных структур, которые установлены с помощью рентгеновской кристаллографии [49]. Обратите внимание на глубокий минимум при 222 нм в КД-спектре а-спирали, который в случае ß-структуры выражен значительно слабее. Для неупорядоченной структуры при этой длине волны КД почти полностью отсутствует. По глубине указанного минимума часто оценивают относительное содержание спиральных участков в белке.
Недавно достигнуты некоторые успехи в предсказании оптического вращения молекул исходя из количественных данных по поляризуемости индивидуальных атомов [50].