Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Биохимическая генетика и синтез нуклеиновых кислот и белков
Транскрипция молекул РНК
Оперон
Копирование генетической информации, заложенной в ДНК, с образованием молекул мРНК представляет собой первый этап в цепи реакций, приводящих к синтезу большого множества жизненно необходимых клетке белков. Поэтому неудивительно, что этот процесс находится под строгим контролем.
Индукция ферментов — одно из тех явлений, которые подвели Жакоба и Моно [36] к модели оперона, описывающей механизм регулирования транскрипции мРНК с ДНК. За эту работу они получили в 1965 г. Нобелевскую премию. В своем окончательном виде [38—40] гипотеза Жакоба — Моно приведена на рис. 15-3. Оперон представляет собой регулируемую группу генов. На рис. 15-3 показан lас-оперок Е. coli. Этот оперон, обнаруживаемый на генетической карте (рис. 15-1) на участке, соответствующем 8 мин, можно, пожалуй, считать наиболее полно исследованной группой генов Е. coli. В его состав входят три структурных гена, кодирующих аминокислотные последовательности ß-галактозидазы (2-ген)1), пермеазы (y-ген) и трансацетилазы с неопределенной функцией (a-ген). Для того чтобы объяснить очевидную синхронность регуляции этих трех генов, Жакоб и Моно предположили, что они образуют одну транскрипционную единицу, которая транскрибируется в виде одного «куска» мРНК.
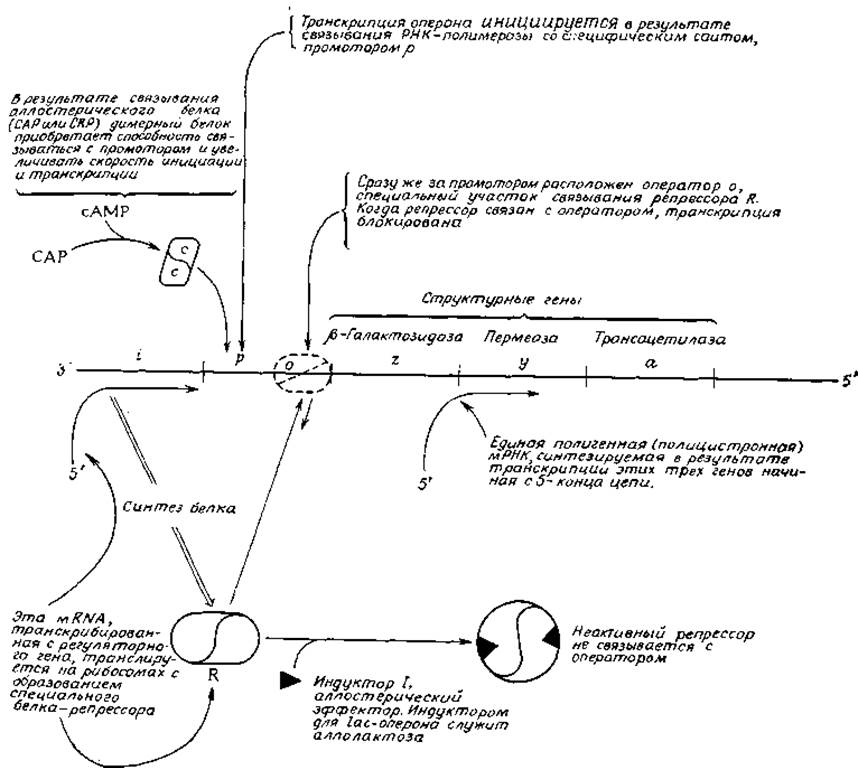
РИС. 15-3. Схематическое изображение lac-оперона Е. coli и его регуляции.
1) В табл. 15-1 н на рис. 15-1 использованы генетические символы lac Z, lac Y и т. д. Однако эти же гены часто обозначают (как это сделано здесь) строчными буквами.
В отличие от других транскрипционных фрагментов оперон находится под контролем специального участка молекулы ДНК, расположенного в начале (на 3'-конце матричной цепи) оперона. Первая часть этого регуляторного участка известна под названием промотора (р). Промотор — это участок ДНК, в котором происходит в самом начале присоединение РНК-полимеразы к ДНК; значения констант связывания, характеризующих присоединение РНК-полимеразы к промоторам, очень велики. Сильное влияние на скорость ассоциации и инициации могут оказывать специальные регуляторные белки. Один из них — сАМР-рецепторный белок (САР)—имеет важное значение для lас-оперона. Он также связывается в промоторном участке (рис. 15-3).
Непосредственно по соседству с промотором расположен оператор (о)—место связывания репрессора (R). Когда оператор свободен, транскрипция запускается и, пройдя операторный участок, доходит до генов, детерминирующих синтез трех указанных выше белков. Но если оператор связан с репрессором, транскрипция блокируется. В то время когда была впервые предложена модель оперона, химическая природа репрессора была неизвестна. Сейчас, однако, уже известно, что в некоторых случаях репрессорами служат белки. Все хорошо изученные репрессоры представляют собой олигомерные белки, способные подвергаться аллостерическим изменениям. Так, lас-репрессор состоит из четырех идентичных субъединиц с мол. весом 37 200, каждая из которых содержит 347 аминокислотных остатков. В каждой субъединице имеется один участок для связывания с оператором и другой (аллостерический) для связывания с эффектором (на рис. 15-3 для простоты вместо четырех субъединиц изображены только две).
lас-Оперон обычно находится в репрессированном состоянии и активируется лишь в присутствии индуктора1). На основании этого Жакоб и Моно постулировали, что с оператором связывается свободный репрессорный белок. В присутствии индуктора конформация репрессора изменяется, и сродство репрессорного белка к операторному участку в результате нарушается. Таким образом, при наличии индуктора оператор не блокирован и гены транскрибируются [36].
1) Длительное время считали, что роль индуктора играет сама лактоза. Однако в действительности оказалось, что эта роль принадлежит аллолактозе (ß-D-Gal-p-(1—>6)D-Glc). В экспериментальных исследованиях чаще всего используют нефизиологические индукторы типа изопропил-β-D-тиогалактозида.
Важную роль в контролировании оперона играет регуляторный ген, детерминирующий синтез репрессорного белка. В случае lас-оперона данный ген (известный под названием i-гена) локализован непосредственно перед lас-опероном (рис. 15-3). Однако регуляторный ген некоторых других оперонов располагается на значительном расстоянии от них. Так, gal-onepoн Е. coli [41] (детерминирующий синтез ферментов, участвующих в метаболизме галактозы) расположен на карте в положении, соответствующем 17 мин, а его регуляторный ген — в положении, соответствующем 61 мин.
Скорость транскрипции регуляторных генов обычно очень низка, но держится на постоянном уровне. Возможно, это объясняется тем, что РНК-полимераза медленнее инициирует синтез цепей РНК на промоторных участках регуляторных генов. Так, в каждой клетке Е. coli в норме содержится всего лишь около 10 молекул lac-репрессорного белка. Поскольку репрессоры имеют очень важное значение для регуляции метаболизма, регуляторные гены представляют чувствительные участки для мутаций. Так, например, мутация регуляторного гена может привести к образованию дефектного репрессора, неспособного более связываться с оператором. В этом случае транскрипция оперона не будет контролироваться и мРНК начнет синтезироваться быстрее. В таком мутантном штамме (в отличие от обычного штамма і+ его обозначают i--) синтез фермента, за который ответственен данный ген, становится «конститутивным», т. е. таким же, как синтез ферментов центральных путей метаболизма. Последние синтезируются регулярно в больших количествах, выйдя из под контроля репрессором. Хорошо известны также и конститутивные операторные мутанты (ос). В этом случае репрессор не может связываться с операторным участком ДНК из-за измененной нуклеотидной последовательности в операторе.
а. Нуклеотидная последовательность регуляторного участка lас-оперона
Замечательное достижение последних лет состоит в установлении нуклеотидной последовательности ДНК Е. coli, в промоторно-операторном участке lас-оперона. Эта последовательность включает конец і-гена и начало 2-гена (рис. 15-4) [39]. Детальное генетическое картирование этой области позволило точно установить нуклеотидную последовательность операторного и промоторного участков. Обратите внимание на расположенный на левом конце ряд кодонов, определяющих последовательность пептида Glu-Ser-Gly-Gln-cтоп. Эта последовательность соответствует С-концевой последовательности репрессора, синтез котоporo кодирует і-ген. На правом конце находятся три кодона, кодирующие формил-Меt-Thr-Меt-N-концевую последовательность ß-галактозидазы, за синтез которой отвечает 2-ген. Была расшифрована также последовательность 63 оснований фрагмента мРНК, считываемого с данного участка lac-оперона [42]. мРНК-транскрипт, как это показано на рис. 15-4, начинается с операторного участка1). Инициаторный кодон гена 2 отстоит от конца на 39 нуклеотидов. Фрагмент с расшифрованной последовательностью нуклеотидов кодирует синтез восьми аминокислот галактозидазы.
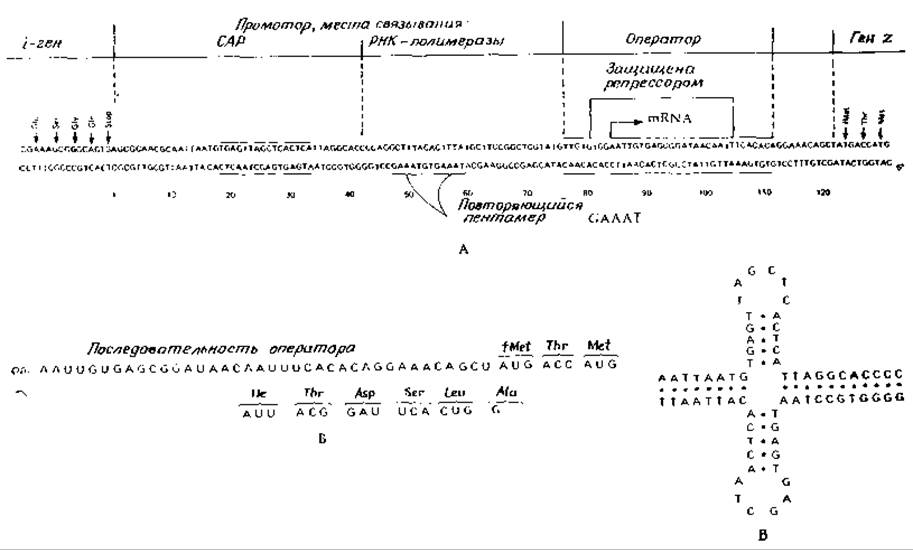
РИС. 15-4. А. Нуклеотидная последовательность lас-промоторно-операторного участка хромосомы Е. coli [43]. Показаны предполагаемые положения гена і, промотора с участками связывания САР- и РНК-полимеразы, оператора и начало гена z (ß-галактозидазы). Обратите внимание на два участка, обладающих вращательной симметрией второго порядка (отмечены горизонтальными линиями) и на повторяющийся пентамер. Б. Нуклеотидная последовательность молекулы мРНК, синтезированной на lас-промоторно-операторном участке мутантного штамма Е. coli с измененным промотором [46, 47]. Место инициации синтеза пептида обозначено символом fMet. Показаны также кодоны для последующих аминокислот N-концевого участка ß-галактозидазы [46]. В. Крестообразная структура, которая может образовываться при выпячивании комплементарных цепей ДНК в САР-связывающем участке. САР — белковый активатор катаболизма (сАМР-связывающий белок).
Нуклеотидная последовательность в операторном участке была установлена [43] путем расщепления ДНК дезоксирибонуклеазой в присутствии репрессорного белка. Будучи связанным, репрессор защищает участок, состоящий из 27 нуклеотидных пар (показано на рисунке). Поразителен тот факт, что центральная часть оператора располагается в участке с вращательной симметрией второго порядка (гл. 2, разд. Г, 11). В результате цепь ДНК оказывается способной образовывать петли, благодаря которым структура ДНК приобретает крестообразную форму (рис. 2-30 и 15-4) [44]. Есть все основания думать, что такая структура может легче связываться с тетрамерным репрессорным белком, чем линейная форма.
Переход линейной двойной спирали в крестообразную структуру требует значительного распрямления спирали, откуда следует, что молекулы с отрицательными супервитками ДНК должны связывать репрессор более прочно, чем ДНК без суперспиральных витков (гл. 2, разд. Г, 9). Экспериментальные данные указывают на то, что развертывание lас-репрессором может осуществляться всего лишь на 40—90°. Таким образом, связывание lас-репрессора сопровождается, по-видимому, слабым нарушением двухспиральной структуры [45]. Тем не менее симметрия второго порядка имеет, по-видимому, важное значение для прочного связывания двух субъединиц симметричного тетрамерного белка.
Возможно также, что молекулы репрессора движутся вдоль цепей ДНК по законам одномерной диффузии и что симметрия операторного участка облегчает его узнавание белком, движущимся к нему в любом из двух направлений.
б. Положительный контроль
Роль важного регуляторного агента в бактериальных клетках играет циклический АМР (сАМР, гл. 7, разд, Д, 8). Примером процесса, опосредованного участием сАМР, может служить катаболитная репрессия. Сущность этого процесса состоит в ингибировании (катаболитом) транскрипции генов, детерминирующих синтез ферментов, необходимых для катаболизма лактозы или других энергетических субстратов, когда в среде присутствует глюкоза — более эффективный источник энергии. Механизм этого процесса не известен, однако установлено, что в присутствии глюкозы концентрация сАМР снижается.
Было показано, что сАМР стимулирует инициацию транскрипции многих оперонов у бактерий. Эта реакция протекает с участием специального связывающего белка, для обозначения которого обычно используют сокращения САР2) или CRP3) [46]. Комплекс САР — сАМР, по-видимому, связывается с промотором на участке, прилегающем к месту связывания РНК-полимеразы. Считают, что непосредственно местом связывания является второй из предполагаемых центров симметрии второго порядка [39] в молекуле ДНК (рис. 15-4). Молекула САР представляет собой димер, состоящий из двух идентичных субъединиц (с мол. весом ~22 000), симметричное расположение которых способствует связыванию в этом участке. Остается еще один важный вопрос: «Каким образом связывание комплекса САР—сАМР приводит к повышению скорости инициации транскрипции мРНК?» К этому вопросу мы вернемся несколько позже, после того, как обсудим свойства РНК-полимераз.
1) В действительности мРНК последовательность которой расшифрована, была выделена из штамма, несущего мутацию в области промотора. Считается, что у Е. coli дикого типа сайт инициации мРНК может быть локализован несколько левее.
2) От английского catabolyte activator protein (белок — активатор катаболизма).
3) Отанглийского сАМР receptor protein (сАМР-рецепториый белок).
Другой тип положительного контроля известен для арабинозного (аrа) оперона (положение, соответствующее 1 мин на хромосомной карте Е. coli). В этом случае индуктор не только вызывает отщепление репрессора от операторного участка, но и превращает его в активатор, который, подобно комплексу САР—сАМР, вызывает более эффективную инициацию транскрипции.
в. Репрессия по принципу обратной связи
Репрессию по принципу обратной связи (гл. 6 разд. Е,2) конечными продуктами биосинтетических реакций можно представить путем простой модификации модели оперона, показанной на рис. 15-3. В таких случаях какой-то продукт, например аминокислота, связывается с анорепрессором, вызывая его аллостерическую модификацию. В этом случае это будет не апорепрессор, а комплекс эффектор — репрессор, связывающийся с опероном и включающий транскрипцию [47]. Роль корепрессоров для оперонов определенных аминокислот, отвечающих, например, за синтез гистидина и валина, играют молекулы аминоацил—тРНК, полученные из этих же аминокислот [48, 49]. В случае оперона триптофана Е. coli транскрипция предотвращается в результате связывания с опероном триптофан-репрессорного комплекса. Был обнаружен, однако, и другой механизм регуляции [50]. Транскрипту гена в мРНК предшествует «лидирующий» участок, состоящий приблизительно из 160 нуклеотидов (последовательность которых известна). Соответствующая последовательность в ДНК содержит два палиндрома, которые играют предположительно роль аттенюаторов (участков затухания синтеза РНК). Если эти места связываются соответствующим белком, то происходит преждевременная терминация цепи и пять последующих генов в опероне не транскрибируются [50].