Химия белка. Структура, свойства, методы исследования - Шендрик А.Н. 2022
Белки кровеносной и мышечной тканей
Белки сократительной системы
Механизм мышечного сокращения - Источники энергии мышечного сокращения
В изолированных мышцах при повторных циклах сокращение-расслабление накапливается молочная кислота и уменьшается содержание гликогена. Сам по себе этот факт может свидетельствовать о том, что необходимая для выполнения работы энергия высвобождается в процессе гликолиза. Исследования показывают, однако, что ингибирование процесса гликолиза не приводит к утрате двигательной функции мышцы. Следовательно, гликолиз не является решающим энергетическим фактором сокращения. Сохраняется сократительная способность мышцы и при блокировании цианидами процесса клеточного дыхания. Из этих фактов следует, что в мышцах содержатся богатые энергией вещества, которые и при ингибировании процесса синтеза АТФ способны поддерживать работоспособность мышц по крайней мере в течении некоторого времени.
Исторически, моменты получения основных экспериментальных данных относительно закономерностей мышечного сокращения и открытия АТФ совпали во времени. Поэтому, одно из первых предположений относительно источника энергии мышечного сокращения в условиях блокирования процессов дыхания и гликолиза сводилось к мысли, что таким источником является исключительно АТФ. Эта гипотеза не нашла, однако, экспериментального подтверждения и была отклонена по следующим основным причинам.
1. Содержание АТФ в мышцах очень мало и не способно обеспечить ее сокращения даже на короткий период времени. Так, например, для поддержания активности мышц млекопитающих необходимо гидролизовать около 10 моля АТФ на 1г мышцы в 1 минуту. Содержание же АТФ в клетках составляет примерно 5∙10-6 моль/г. Этого количества АТФ достаточно не более чем на 0.5 с работы мышцы
2. Измерениями содержания АТФ в мышце до и после сокращения не было выявлено уменьшения АТФ или увеличения АДФ по завершению цикла сокращения.
Дальнейшими исследованиями было установлено, что в мышцах содержатся достаточно большие запасы фосфагенов. Наиболее важными из них являются креатинфосфат (обнаружен у большинства позвоночных) и аргининфосфат (у беспозвоночных).
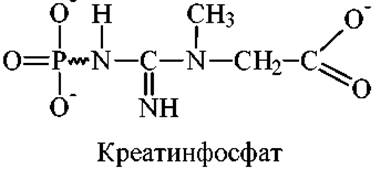
Креатинфосфат представляет собой производное гуанидина, в молекуле которого атом фосфора соединен непосредственно с азотом. При гидролитическом отщеплении фосфатной группы выделяется большое количество энергии:
Креатинфосфат + АДФ Креатин + АТФ, ∆G0 = -12 кДж/моль
Реакция катализируется ферментом - креатинкиназой. При физиологическом значении рН = 6 равновесие смещено влево. Поэтому при единичных сокращениях мышцы величина квазиравновесной концентрации АТФ практически не изменяется, поскольку имеется канал быстрого восстановления АТФ за счет креатинфосфата, содержание которого превышает концентрацию АТФ в мышцах в 4-5 раз. Креатинкиназный путь ресинтеза АТФ является быстрым и максимально эффективным источником АТФ. Если стимулировать сокращение мышцы в течение более длительного времени в отсутствии гликолиза или дыхания, то содержание креатинфосфата уменьшается. Это, в свою очередь, ведет к уменьшению концентрации АТФ и утрате, в конечном итоге, сократительной способности мышцы.
Важная роль креатинфосфата, как резервуара АТФ, подтверждена также в экспериментах с ингибиторами (Кейн с соавт., 1962г). Так, креатинфосфокиназную активность интактных мышц можно полностью подавить 2,4-динитрофторбензолом (реактивом Сенгера). В мышцах, отравленных этим ингибитором, наблюдается очень быстрое уменьшение концентрации АТФ при сокращениях. Это следствие того, что канал образования АТФ из креатинфосфата заблокирован 2,4-динитрофторбензолом.
Имеются сведения о том, что креатинфосфат используется в мышце, особенно сердечной, не только как депо легкомобилизуемых макроэргических фосфатных связей, но и транспортирует богатые макроэргичесикими фосфатными связями продукты окислительного фосфорилирования. Предложена схема такого переноса из митохондрий в цитоплазму клеток миокарда. Согласно этой схеме, АТФ из матрикса митохондрий переносится через их внутреннюю мембрану с участием специфической АТФ-АДФ-транслоказы на активный центр митохондриального изофермента креатинкиназы. Последний расположен на внешней стороне внутренней мембраны митохондрий. В межмембранном пространстве, в присутствии ионов магния, и при наличии в среде креатина образуется тройной фермент-субстратный комплекс: креатин-креатинкиназа-АТФ-Mg2+. Этот комплекс распадается затем с образованием креатинфосфата и АДФ-Mg2+. Креатинфосфат диффундирует в цитоплазму, где используется в миофибриллярной креатинкиназной реакции для рефосфорилирования. Предполагается, что аналогичный путь транспорта энергии из митохондрий в миофибриллы реализуется и в скелетной мускулатуре.
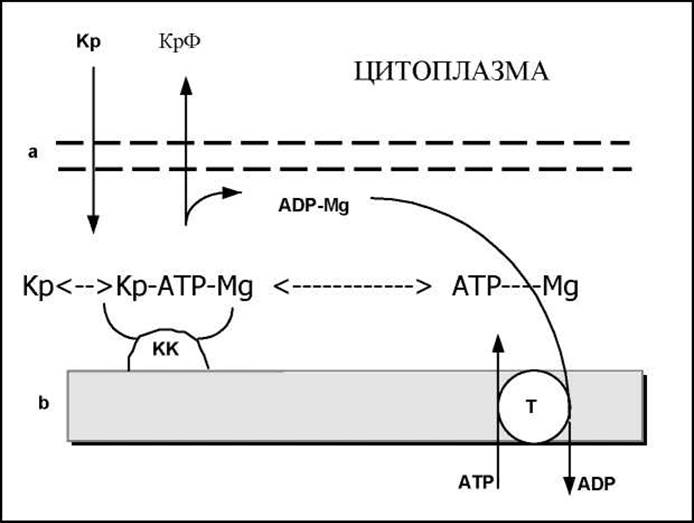
Процесс переноса энергии из митохондрий в цитоплазму клетки миокарда (по Саксу и др.) схематически представлен на рис.
Где а, b - наружная и внутренняя мембраны соответственно; Кр - креатин; КрФ - креатин фосфат; КК - креатинкиназа; Т - Транс-локаза
Процессы рефосфорилирования АДФ до АТФ и образования креатинфосфата поддерживаются во всех мышцах гликолизом и дыхательной активностью. Однако, вклад гликолиза и дыхания в разных мышцах различен. В высокоактивных, или красных мышцах (цвет обусловлен высоким содержанием миоглобина и цитохромов) основным источником является окислительное фосфорилирование в митохондриях. К этим мышцам относятся летательные мышцы птиц и мышцы конечностей млекопитающих. В малоактивных, или белых скелетных мышцах с малым содержанием миоглобина и цитохромов основной источник энергии гликолиз.