Принципы структурной организации белков - Г. Шульц 1982
Взаимодействия, определяющие структуру белка
Молекулярная упаковка
Энергия водородных связей используется эффективно. После общего рассмотрения невалентных сил, способствующих стабильности свернутой полипептидной цепи, попытаемся оценить их действительную эффективность. Исследования белковых структур показывают, что около 90% всех внутренних полярных групп образуют водородные связи [17]. Это согласуется с большим содержанием вторичной структуры, наблюдающимся в белках (гл. 5), и показывает, что фактически именно на ее образование расходуется почти вся возможная энергия водородных связей.
Высокая плотность упаковки согласуется со значительной ролью дисперсионных сил. Об эффективности участия дисперсионных сил можно судить по плотности упаковки внутри белка, поскольку плотность упаковки отражает количество имеющихся контактов. Плотность упаковки определяется как отношение собственного объема молекулы (объем, заключенный внутри вандерваальсовой оболочки молекулы, который в свою очередь определяется вандерваальсовыми радиусами атомов на поверхности) к объему, реально занимаемому молекулой в пространстве. В кристаллах малых молекул реально занимаемый объем задается элементарной ячейкой, а объем молекулы внутри вандерваальсовой оболочки можно определить по координатам атомов в кристалле и их вандерваальсовым радиусам.
Чтобы определить общую плотность упаковки молекулы белка, вандерваальсов объем вытянутой цепи следует отнести к объему свернутой цепи, т. е. к объему молекулы нативного белка. Однако целесообразно вначале определить локальную плотность упаковки, которая в белке может варьировать в зависимости от определенных структурных особенностей. Интегрируя по всем локальным плотностям упаковки, можно затем получить среднюю плотность упаковки белка.
Полиэдр Вороного позволяет определить локальную плотность упаковки. Локальные плотности упаковки были впервые определены Ричардсом [63]. Для этой цели полипептидная цепь была разделена на небольшие атомные группы, такие, как метил, метилен, амид, гидроксил, и другие, содержащие один более тяжелый атом и до трех атомов водорода. Ароматические циклы и гуанидиниевая группа Arg также рассматривались как отдельные группы. Из рентгеноструктурных данных были определены центры этих групп. Затем пространство было разбито на полиэдры Вороного, как показано на рис. 3.4. Локальная плотность упаковки при данном центре есть отношение объема атомной группы к соответствующему занимаемому объему, т. е. к объему окружающего полиэдра Вороного. Чтобы избежать ошибок, связанных с учетом поверхности, в рассмотрение был включен монослой молекул Н2О, находящихся на поверхности. Таким образом, в полный полиэдр Вороного были включены только атомы белка, но не фактические молекулы Н2О. Это поясняется на рис. 3.4, где А — центр атома белка, а В, С и D — центры фактических молекул Н2О (см. также рис. 1.9). Эта процедура позволяет попутно определить площадь молекулярной поверхности. Лучше всего воспользоваться тем монослоем поверхностных молекул воды, который содержит их максимальное количество. Однако поскольку определить такой оптимальный слой трудно. то выбранный слой обычно содержит меньшее по сравнению с оптимальным число молекул воды. Вводимая за счет этого ошибка невелика.
Белки упакованы так же плотно, как хорошие молекулярные кристаллы. Наблюдаемые локальные плотности упаковки в белках варьируют от 0,68 до 0,82. Низкая плотность найдена в активных центрах [63, 64], что подтверждает предположение о подвижности активных центров. Высокую плотность имеют гидрофобные ядра в центре белка*. Средняя плотность упаковки белка составляет около 0,75 (плотность упаковки правильных твердых сферических тел составляет 0,74). Для кристаллов малых молекул, связанных вандерваальсовыми силами, характерны значения от 0,70 до 0,78, в среднем 0,74. Стекла, масла или исключительно мягкие вандерваальсовы кристаллы (или некоторые кристаллы, построенные за счет направленных связей, например водородных связей обычного льда, рис. 3.2) имеют плотность упаковки ниже 0,70 и даже ниже 0,60. Таким образом, белки действительно упакованы столь же плотно, как малые молекулы в молекулярных кристаллах; естественно, что для них должна играть большую роль дисперсионная энергия.
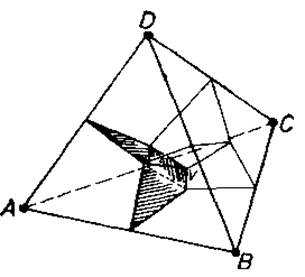
Рис. 3.4. Разбиение пространства на полиэдры вокруг атомов.
Для данного набора атомов пространство можно разбить на неправильные полиэдры с атомами в их вершинах; один из таких полиэдров приведен на этом рисунке. Как показал Вороной (1908), четыре биссекторные плоскости между каждой парой атомов пересекаются в одной точке — вершине V. Эти плоскости делят тетраэдр на четыре неравные части. Часть, прилежащая к атому А, выделена заштрихованными плоскостями. Данный атом А может принадлежать произвольному числу тетраэдров. Чтобы определить общий полиэдр вокруг А, приведенное построение необходимо выполнить для всех таких тетраэдров и все части, прилежащие к А, нужно сложить. После повторения этой процедуры для всех атомов (атомы на поверхности набора исключаются) пространство полностью разделено. Плотность упаковки данного атома определяется как отношение объема внутри вандерваальсовой оболочки к объему окружающего полиэдра. Ричардс [63] перенес эту процедуру на белковые молекулы, которые содержат атомы нескольких типов с разными вандерваальсовыми радиусами. В этом случае процедура Вороного, которая дает точные результаты для одинаковых атомов, в своем первоначальном виде неприменима. В ее модифицированной форме, дающей удовлетворительные результаты, биссекторные плоскости проводятся через точки, которые делят расстояние между атомами, например между А и В, пропорционально соотношению вандерваальсовых радиусов А к В или, если А и В связаны ковалентно, в соответствии с отношением их ковалентных радиусов. Вследствие таких смещений четыре плоскости уже не пересекаются в точке V, а образуют небольшой тетраэдр вблизи V. Относительный объем этого «ошибочного» тетраэдра всего около 1%, и им можно пренебречь, если отношение радиусов не превышает 1,5. Чтобы эти отношения оставались на уровне 1,0, Ричардс ввел в атомные группы все (малые) Н-атомы.
* Плотность упаковки нельзя смешивать с электронной плотностью или с плотностью массы. Гидрофобные ядра, как и все углеводороды, имеют значительно более низкую плотность массы, чем полярные участки.
Наблюдаемые плотности упаковки показывают, что метафору Козмана о капле масла не следует понимать буквально. Внутренняя часть белка более похожа на кристалл. Это подтверждается также слабой сжимаемостью белков по сравнению с маслом [65, 66]: kмаслo/kбелок ≈ 20.
Упаковка сохраняется в процессе эволюции. Концепция плотной упаковки белка и, следовательно, связанных с ней стерических ограничений, должна быть согласована с образованием большого числа направленных водородных связей практически всеми полярными группами внутри его молекулы. Напомним в связи с этим, что в обычном льде именно из-за образования линейных водородных связей плотность упаковки составляет всего 0,58. Высокая структурная оптимизация в белке подтверждает предположение о том, что эволюционные изменения гораздо реже касались внутренней части белка, чем его поверхности. Кроме того, имеется определенная тенденция ко взаимной компенсации внутренних изменений [67], например изменение Ile → Val может сопровождаться соседним изменением Gly → Ala, которое восстанавливает потерянную метальную группу.
Плотность упаковки может служить критерием в предсказании структуры. Плотность упаковки предоставляет возможность независимо проверить результаты расчетного моделирования свертывания цепи (разд. 8.6) и предсказания структур по известным аминокислотным последовательностям путем сравнения с филогенетически связанными белками (гл. 9). Этот критерий не сводится только к сравнению со средними плотностями упаковки. Объем, занимаемый боковыми цепями (сумма объемов всех составляющих данную цепь атомных групп), специфичен для каждой боковой цепи [63, 68] и варьирует всего лишь приблизительно на 5%. Поэтому в качестве еще одного критерия в предсказании белковой структуры можно воспользоваться объемом, занимаемым каждой боковой цепью.