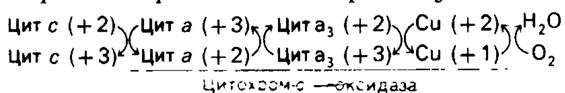БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 14. ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ
14.4. Флавин, железосерные комплексы, хинон и гемовые группы переносят электроны от NADH к O2
Электроны переносятся от NADH к O2 через ряд переносчиков: флавины, железосерные комплексы, хиноны и гемы (рис. 14.4). Эти переносчики электронов, за исключением хинонов, являются простети- ческими группами белков. Первая реакция состоит в окислении под действием NАDН-Q- редуктазы (называемой также NАDН-дегидрогеназой),ферментом, состоящим по меньшей мере из шестнадцати полипептидных цепей. Два электрона переносятся от NADH на флавинмонопуклеотид (FMN), простетическую группу фермента, с образованием ее восстановленной формы FMNH2
NADH + Н+ + FMN → FMNH2+ NAD+.
Рис. 14.4. Последовательность переносчиков электронов в дыхательном ансамбле. Протоны выбрасываются тремя комплексами, которые на рисунке закрашены

C FMNH2 электроны переносятся затем на ряд железо-серных комплексов (сокращенно Fe-S), играющих роль второй нро- стетической группы в молекуле NADH-Q- редуктазы. Железо в этих соединениях это негемовое железо, в связи с чем железосерные белки называют также не гемовыми железопротеинами, Новые исследования показали, что Fe-S-комплексы играют важную роль в широком круге окислительно-восстановительных реакций в биологических системах. Известны три вида Fe-S-центров (рис. 14.5). В простейшем случае единственный атом железа тетраэдрически координирован с сульфгидрильными группами четырех цистеиновых остатков белка. Второй вид комплексов (обозначен как Fe2-S2) содержит 2 атома железа и два неорганических дисульфида, присоединенных к четырем цистеиновым остаткам. В комплексах третьего вида (Fe4-S4) содержится четыре атома железа, четыре неорганических сульфида и четыре остатка цистеина. Атом железа в этих комплексах может присутствовать в восстановленном (Fe2+) или окисленном (Fe3+) состоянии. NADH-Q-редуктаза содержит второй (Fe2-S2) и третий (Fe4 -S4) тины комплексов.
От железосерных центров NADH-Q-peдуктазы электроны переносятся далее на кофермент Q (сокращенно Q).
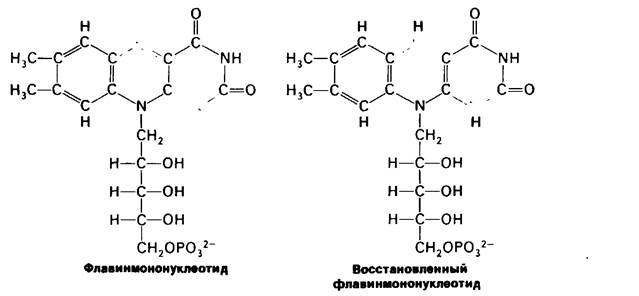

Рис. 14.5. Молекулярные модели Fe-S- комплексов: А) Fe-S-центр; Б) Fe2-S2-HeHTp; В) Fe4-S4- центр. Атомы железа окрашены красным цветом, атомы серы цистеина - желтым, атомы неорганической серы- зеленым
Кофермент Q-хиноновое производное с длинным изопреноидным хвостом. Его называют также убихиноном из-за его повсеместного распространения (ubiquitous) в биологических сисгемах. Число изопреновых единиц в коферменте Q зависит от вида живых организмов. У млекопитающих его наиболее распространенная форма содержит десять изопреновых единиц и обозначается как Q10.
Изопреноидный хвост обусловливает высокую неполярность Q, которая способствует его быстрой диффузии в углеводородной фазе внутренней митохондриальной мембраны. Кофермент Q-единственный переносчик электронов в дыхательной цепи, который не связан прочно с белком и не присоединен к нему ковалентно. Кофермент Q действительно служит высокомобильным переносчиком электронов между флавопротеинами и цитохромами цепи переноса электронов.
Напомним, что FADH2 образуется в цикле трикарбоновых кислот при окислении сукцината в фумарат сукцинат-дегидрогеназой. Сукцинат-дегидрогеназа - один из двух компонентов сукцинат-Q-редуктазного комплекса; второй компонент Fe-S-белок. Этот комплекс, подобно NADH-Q-редуктазе, является интегральным компонентом внутренней митохондриальной мембраны. Обладающие высоким потенциалом электроны FADH2в сукцинат-дегидрогеназном компоненте переносятся затем на Fe-S-центры комплекса и далее на кофермент Q для включения в цепь переноса электронов. Подобно этому, глицеролфосфат-дегидрогеназа (разд. 14.9) и дегидрогеназа СоА-эфиров жирных кислот (разд. 17.8) переносят свои электроны с высоким потенциалом на кофермент Q с образованием его восстановленной формы QH2.
Переносчиками электронов между QH2 и 02 помимо одного Fe-S-белка служат все цитохромы. Центральную роль цитохромов в дыхании открыл в 1925 г. Дэвид Кейлин (David Keilin). Цитохром-это переносящий электроны белок, молекула которого содержит в качестве простетической группы гем. В ходе переноса электронов атом железа находится то в восстановленной ферроформе (+ 2), то в окисленной ферриформе (+ 3). Группа гема, подобно Fe-S-центру, переносит только один электрон в отличие от NADH, флавина и кофермента Q, переносящих по два электрона. Таким образом, молекула QH2, восстановленной формы хинона, переносит свои два обладающие высоким потенциалом электрона на две молекулы цитохрома Ь, следующего члена цепи переноса электронов. В качестве промежуточного продукта при этих реакциях переноса электронов, возможно, образуется свободный радикал семихинона (обозначенный QH •).
В цепи переноса электронов на участке от QH2до О2 располагаются пять цитохромов. Цитохромы δ и с1 наряду с Fe-S-белком являются компонентами QH2-цитохром-с - редуктазного комплекса. Цитохром с переносит электроны от этого комплекса к цитохром-с-оксидазному комплексу, содержащему цитохромы а и а3. Окислительно-восстановительный потенциал (сродство к электронам, или окислительная способность) этих цитохромов возрастает в последовательности
QН2 → Цитохром δ → FеS → Цитохром c1→ Цитохром с → Цитохром а → Цитохром а3 → O2.
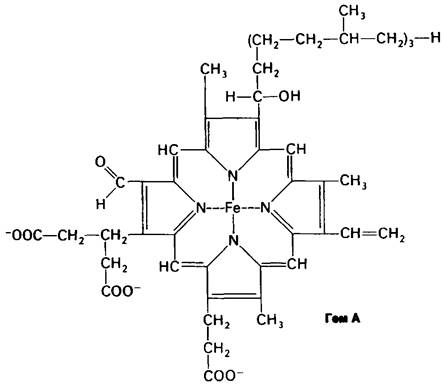
Указанные цитохромы различаются по структуре и свойствам. Простетической группой цитохромов Ь, с1 и с служит железосодержащий протопорфирин IX, называемый обычно гемом, который является простетической группой также в миоглобине и гемоглобине. В цитохроме δ гем не связан ковалентно с белком, тогда как в цитохромах с и с1 он ковалентно присоединяется к белку при помощи тиоэфирных связей (рис. 14.6). Эти связи образуются путем присоединения сульфгидрильных групп двух цистеиновых остатков к винильным группам гема.
Рис. 14.6. Гем в цитохромах с и с1 ковалентно присоединен к двум цистеиновым боковым цепям
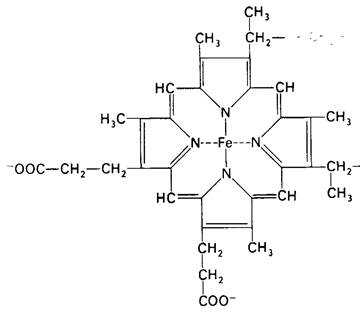
Цитохромы а и а3 имеют иную железо- порфириновую простетическую группу, называемую гемом а. Он отличается от гема цитохромов с и c1 наличием формильной группы вместо одной из метальных групп и углеводородной цепи вместо одной из винильных групп. Цитохромы а и а3 являются конечными звеньями дыхательной цепи. Они существуют в виде комплекса, иногда называемого цитохромоксидазой.
QН2-цитохром-с-редуктазный комплекс переносит электроны от QН2 к цитохрому с, водорастворимому периферическому мембранному белку (разд. 14.15-14.17).
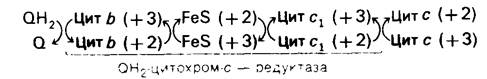
Затем восстановленный цитохром с переносит свои электроны на цитохром-с-оксидазный комплекс. Роль цитохрома с аналогична роли кофермента Q: он служит мобильным переносчиком электронов между различными комплексами в дыхательной цепи. Электроны переносятся к a-цитохромному компоненту комплекса и затем к цитохрому а3, содержащему медь. При транспорте электронов от цитохрома а3 к молекулярному кислороду атом меди существует то в окисленной (+2), то в восстановленной (+1) форме. Образование воды-процесс, связанный с переносом четырех электронов, тогда как группы гема являются переносчиками одного электрона. Каким образом четыре электрона «сходятся» для восстановления молекулы O2, пока не установлено:
O2 + 4Н+ + 4е- → 2Н2O.