БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 14. ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ
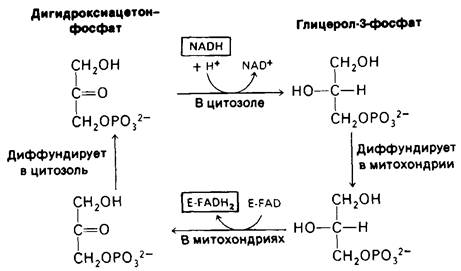
14.9. Электроны от цитоплазматического NADH поступают в митохондрии при помощи глицерофосфатного челночного механизма
Интактные митохондрии непроницаемы для NADH и NAD+. Как же тогда происходит окисление цитоплазматического NADH дыхательной цепью? NADH образуется в процессе гликолиза при окислении глицеральдегид-3-фосфата. Чтобы гликолиз продолжал функционировать, должно происходить генерирование NAD+. Решение этой проблемы состоит в том, что через митохондриальную мембрану переносится не сам NADH, а отдаваемые им электроны. Одним из переносчиков является глицерол-3-фосфат, который легко проходит через наружную митохондриальную мембрану. Первый шаг в этом челночном механизме (рис. 14.11), перенос электронов от NADH на дигидроксиацетонфосфат с образованием глицерол-3-фосфата. Эта реакция, катализируемая глицерол-3-фосфат - дегидрогеназой, протекает в цитозоле. Глицерол-3-фосфат поступает затем в митохондрии, где он снова окисляется в дигидроксиацетонфосфат при участии FAD- простетической группы дегидрогеназы, которая связана с внутренней митохондриальной мембраной. FAD-зависимая глицерол- дегидрогеназа митохондрий отличается от NAD+ зависимой глицерол-дегидрогеназы цитозоля. Образовавшийся при окислении глицерол-3-фосфата дигидроксиацетонфосфат далее диффундирует из митохондрий в цитозоль, завершая челночный процесс.
Рис. 14.11. Глицеролфосфатный челночный механизм
Суммарная реакция может быть изображена следующим образом:

Восстановленный флавин внутри митохондрий переносит свои электроны на дыхательную цепь на уровне кофермента Q. Таким образом, при окислении дыхательной цепью NADH, который переносится глицеролфосфатным челночным механизмом, образуются две, а не три молекулы АТР. На первый взгляд может показаться, что в каждом цикле этого процесса одна молекула АТР теряется. Такой низкий выход связан с предпочтительным использованием в митохондриях FAD, а не NAD+ в качестве акцептора электронов в реакции, катализируемой глицерол-3-фосфат - дегидрогеназой. Использование FAD делает возможным перенос электронов в митохондрии от цитоплазматического NADH против градиента концентрации NADH. «Цена» такого транспорта-одна молекула АТР на два электрона. Глицеролфосфатный челночный механизм играет особенно важную роль в летательных мышцах насекомых.
В сердце и печени электроны транспортируются в митохондрии от цитоплазматического NADH благодаря малат-аспартатному челночному механизму, который опосредуется двумя мембранными переносчиками и четырьмя ферментами. В цитозоле происходит перенос электронов от NADH на малат, который проходит сквозь внутреннюю митохондриальную мембрану и затем вновь окисляется с образованием NADH в митохондриальном матриксе. Оксалоацетат не проходит легко через вну-
треннюю митохондриальную мембрану и должен превратиться в результате реакции трансаминирования в аспартат, способный проходить сквозь данный барьер. Суммарная реакция малат-аспартатного челночного механизма описывается следующим уравнением:
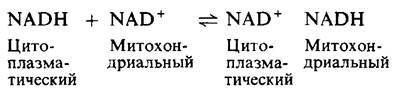
Этот механизм в противоположность глицеролфосфатному челночному механизму характеризуется легкой обратимостью. Следовательно, NАDН может поступать в митохондрии по малат-аспартатному челночному механизму только при условии, что отношение [NADH]/[НАD+] выше в цитозоле, чем в митохондриальном матриксе. При переносе электронов от NADH к митохондриальной дыхательной цепи с помощью данного механизма не происходит поглощения энергии, и на каждую транспортируемую молекулу NАDН синтезируются три молекулы АТР.
14.10. Поступление ADP в митохондрии требует выхода АТР
АТР и ADP не могут диффундировать свободно через внутреннюю митохондриальную мембрану. Переход этих высокозаряженных молекул через этот барьер оказывается возможным благодаря наличию специфического переносчика. Интересная особенность переноса-сопряжение токов АТР и ADP. ADP поступает в митохондриальный матрикс только при условии выхода АТР и наоборот. Этот сопряженный поток АТР и ADP представляет собою пример облегченной обменной диффузии. Он опосредуется ATP-ADP—транслоказой, димером, состоящим из идентичных субъединиц с мол. массой 29 кДа каждая. Транслоказа имеется в большом количестве во внутренней митохондриальной мембране, составляя около 6% общего белка. На цитоплазматическую сторону мембраны транспортируется предпочтительно ADP. Именно этим отчасти и объясняется тот факт, что соотношение [ATP]/[ADP] [Pi] на той стороне мембраны, которая обращена к цитоплазме, в 10 раз выше, чем на стороне, обращенной к матриксу. Сопряженный транспорт АТР и ADP транслоказой, вероятно, индуцируется протонным градиентом через внутреннюю митохондриальную мембрану. ATP-ADP—транслоказа специфически ингибируется очень низкими концентрациями атрактилозида, растительного гликозида. или бонгкрековой кислоты, антибиотика из плесени. Вскоре после добавления этих ингибиторов окислительное фосфорилирование прекращается, поскольку поступления ADP внутрь митохондрии больше не происходит.
14.11. Митохондрии содержат многочисленные транспортные системы для ионов и метаболитов
ATP-ADP—транслоказа-это только одна из многих транспортных систем митохондрий. Внутренняя митохондриальная мембрана содержит целый ряд переносчиков для ионов и заряженных метаболитов. Например, переносчик дикарбоновых кислот опосредует облегченную обменную диффузию малата, сукцината, фумарата и Рi. Переносчики трикарбоновых кислот обеспечивают обмен ОН- на Рi. Пируват цитозоля поступает в митохондриальный матрикс в обмен на ОН- с помощью переносчика пирувата. Взаимный обмен глутамата и аспартата осуществляет переносчик глутамата, способный также переносить ОН-. Митохондрии содержат и систему транспорта ионов кальцин. Непосредственным источником свободной энергии для накопления Са2+ в митохондриальном матриксе является не АТР, а протонодвижущая сила, генерируемая переносом электронов.