БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 15. ПЕНТОЗОФОСФАТНЫЙ ПУТЬ И ГЛЮКОНЕОГЕНЕЗ
15.17. Оксалоацетат переходит по челночному механизму в цитозоль и превращается в фосфоенолпируват
Пируват-карбоксилаза - митохондриальный фермент, тогда как другие ферменты глюконеогенеза находятся в цитоплазме. Оксалоацетат, продукт пируват-карбоксилазной реакции, переносится через митохондриальную мембрану в форме малата. Его восстановление в малат происходит в митохондриях под действием NADH-зависимой малат-дегидрогеназы. Образующийся при этом малат транспортируется переносчиком через митохондриальную мембрану и вновь окисляется в оксалоацетат NAD+-зависимой малат-дегидрогеназой цитозоля.
Оксалоацетат подвергается в цитозоле одновременно декарбоксилироваиию и фосфорилированию под действием фосфоенол- пируват-карбоксикиназы.
На этой стадии происходит отделение СО2, присоединившегося к пирувату под действием прируват-карбоксилазы. Реакция фосфорилирования становится энергетически возможной благодаря одновременному декарбоксилироваиию. Процессы де- карбоксилирорания часто приводят в движение реакции, которые в ином случае были бы высокоэндергоническими. Мы вновь встретимся с этим механизмом в разделе, посвященном синтезу жирных кислот.
15.18. Шесть высокоэнергетических фосфатных связей расходуются при синтезе глюкозы из пирувата
Стехиометрия глюконеогенеза описывается уравнением
2 Пируват + 4 АТР + 2GTP + 2NADH + 2Н2О → Глюкоза + 4ADP + 2GDP + 6 Рi + 2NAD+
∆G0 = - 9 ккал/моль.
Стехиометрия обращения гликолиза носит иной характер:
2 Пируват + 2АТР + 2NADH + 2Н2O →Глюкоза + 2АDР + 2Рi + 2NAD+
∆G0 = + 20 ккал/моль.
Заметим, что для синтеза глюкозы из пирувата путем глюконеогенеза используется шесть высокоэнергетических фосфатных связей, тогда как в процессе превращения глюкозы в пируват при гликолизе образуются только две молекулы АТР. Таким образом, избыточная «цена» глюконеогенеза равна четырем высокоэнергетическим фосфатным связям в расчете на одну молекулу глюкозы, синтезируемой из пирувата. Для превращения энергетически неблагоприятного процесса (обращение гликолиза, ∆G0' = + 20 ккал/моль) в энергетически благопрятный (глюконеогенез, ∆G0 = - 9 ккал/моль) требуются четыре лишние высокоэнергетические фосфатные связи. Рассматривая это энергетическое различие между гликолизом и глюконеогенезом под другим углом зрения, напомним, что вклад одного эквивалента АТР изменяет константу равновесия реакции примерно в 108 раз (разд. 11.7). Следовательно, вклад четырех дополнительных высокоэнергетических связей при глюконеогенезе изменяет эту константу в 1032 раз, что делает превращение пирувата в глюкозу термодинамически выгодным.
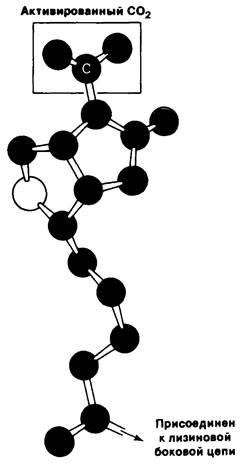
Рис. 15.6. Молекулярная модель карбоксибиотина
15.19. Глюконеогенез и гликолиз регулируются реципрокно
Глюконеогенез и гликолиз координируются таким образом, что, когда активность одного из этих путей находится на относительно низком уровне, другой путь является высокоактивным. В случае одновременной высокой активности обеих последовательностей реакций происходил бы гидролиз четырех ~ Р (два АТР + два GTP) на каждый цикл реакции. В условиях, существующих в клетке, и гликолиз, и глюконеогенез представляют собою высокоэкзергонические процессы, так что термодинамических барьеров для осуществления таких циклов нет. Тот факт, что активность этих двух процессов никогда не достигает высокого уровня одновременно, обусловливается скорее всего соответствующей регуляцией отдельных ферментов каждого процесса. Например, АМР стимулирует фосфофруктокиназу (разд. 12.9), но ингибирует фруктозо-1,6-бисфосфатазу. Цитрат оказывает на эти ферменты противоположное действие. Следовательно, фосфорилирование фруктозо-6- фосфата, этап, лимитирующий скорость гликолиза, усиливается при низком энергетическом заряде клетки. Напротив, при высоком энергетическом заряде и избытке промежуточных продуктов цикла трикарбоновых кислот происходит гидролиз фруктозо-1 ,6-бисфосфата и стимулируется глюконеогенез. Пируваткиназа (разд. 12.7) и пируват-карбоксилаза (разд. 15.16) также регулируются реципрокно. Фруктозо-1,6- бисфосфат стимулирует, а АТР ингибирует пируваткиназу, тогда как пируват-карбоксилаза стимулируется ацетил-СоА и подавляется ADP. Таким образом, обогащение клеток печени топливными молекулами и АТР благоприятствует превращению пирувата в фосфоенолпируват и глюконеогенезу.