Альгологія - Костіков І.Ю. - 2009-2013
Розділ 2. Водорості у системі органічного світу
Надцарства органічного світу
За фенотипічними системами, органічний світ поділяється на два надцарства - Procaryota - прокаріоти, та Eucaryota - евкаріоти. До прокаріот відносяться всі бактерії та один з відділів нижчих рослин - синьозелені водорості. Всі інші організми - тварини, гриби та рослини та як нижчі, так і вищі, належать до евкаріот.
Відміни між прокаріотами та евкаріотами на геномному рівні полягають у тому, що прокаріотична клітина є системою, яка містить лише один геном, зосереджений у нуклеоїді, тобто є моногеномною. Евкаріотична клітина є системою з кількома (двома, трьома або навіть чотирма) неспорідненими геномами, тобто є полігеномною. Геноми евкаріотичної клітини представлені: а) ядерним геномом, зосередженим у ядрі і представленим ядерною ДНК; б) мітохондріальним геномом, зосередженим в мітохондрії і представленим мітохондріальною ДНК; в) пластидним (хлоропластним) геномом, що розташовується в пластиді, і представлений хлоропластною ДНК (генофором); г) геномом нуклеоморфу, який виявлений лише у кількох відділах водоростей у надзвичайно цікавій ядроподібній структурі, розташованій між оболонкою пластиди та особливою клітинною системою - хлоропластною ендоплазматичною сіткою. В нуклеоморфі виявлена власна, нуклеоморфна ДНК. Двогеномні клітини, в яких представлено ядерний та мітохондріальний геноми, характерні для грибів та тварин; тригеномні з ядерним, мітохондріальним та пластидним геномами - для майже всіх евкаріотичних рослин; чотиригеномні з ядерним, мітохондріальним, пластидним та нуклеоморфними геномами виявлені у хлорарахніофітових та криптофітових водоростей, та у деяких видів з відділу Dinophyta.
Клітини прокаріот та евкаріот на фенотипічному рівні схожі за наявністю ДНК та білок-синтезуючого апарату, представленого рибосомами, за наявністю клітинних мембран, зокрема - зовнішньої мембрани, яка називається плазмалема, ферментних комплексів, що забезпечують процеси реплікації, транскрипції та трансляції та синтез АТФ. До складу клітин прокаріот та евкаріот входять білки, жири, вуглеводи, нуклеінові кислоти, мінеральні речовини та вода.
Відміни у будові клітин прокаріот та евкаріот пов'язані, в першу чергу, з тим, що ДНК прокаріот не має хромосомної організації і не пов'язана з білками-гістонами, гени організовані у групи, які називають оперонами, ДНК нуклеоїду прикріплюється до плазмалеми і процес поділу нуклеоїду зумовлений не роботою веретена, а роботою плазмалеми. Рибосоми у прокаріот менші та легкіші від рибосом цитоплазми евкаріот (умовна вага прокаріотичних рибосом становить 70S, тоді як евкаріотичних - 80S). Прокаріоти не здатні до фаго- та піноцитозу, не мають морфологічно оформленного ядра, мітохондрій, пластид, ендоплазматичної сітки, комплексу Гольджі, лізосом, пероксисом, а також органел, що побудовані з мікротрубочок - джгутиків, базальних тіл джгутиків, клітинного центру з центриолями, джгутикових коренів, веретена поділу. У прокаріот відсутні мітоз, мейоз, статевий процес, а обмін генетичною інформацією здійснюється парасексуально - шляхом трансформацій та кон'югацій. Прокаріотичні клітини за розмірами значно менші від евкаріотичних - їх середній діаметр становить біля 0.5-2 мкм, тоді як у евкаріот - 5-20 мкм (рис.1.1).
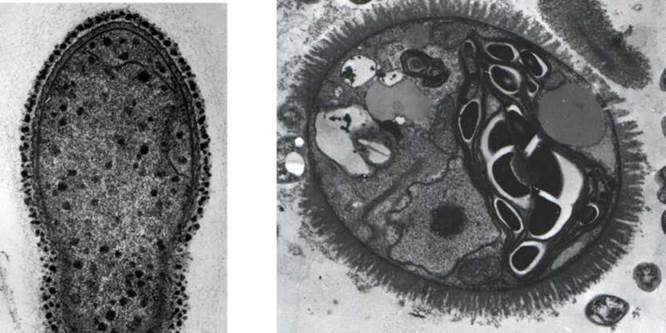
Рис. 1.1. Прокаріотична бактеріальна та евкаріотична водоростева клітини (ліворуч та праворуч, відповідно). У прокаріотичної клітини помітно оболонку з електронно-щільним шаром муреїну, плазмалему, світлу зону в центрі клітини, де розташовується ДНК, включення. У евкаріотичної клітини чітко розрізняються оболонка зі слизовими фібрилами, ядро з ядерцем, мітохондрії, хлоропласт з піреноїдом, вакуоля з клітинним соком, включення. У нижньому правому куті на фотографії представлені також дві бактеріальні клітини, які дають уявлення про відміни прокаріот та евкаріот за розмірами. Фотографії А. Массальського. (додати збільшення або шкалу).
За палеонтологічними даними, прокаріоти на нашій планеті виникли біля 3.2 млрд років тому, тоді як евкаріоти є набагато молодшими - їх вік складає лише біля 1.6 млрд. років. Сьогодні в межах надцарства прокаріот відомо біля 40 тис. видів, тоді як у надцарстві евкаріот - більш від 1.7 млн. видів.
Систематична структура надцарства Procaryota
За молекулярними даними, які добре узгоджуються з фенотипічними ознаками, до надцарства Procaryota входять два царства: Archaebacteria - архебактерії та Eubacteria - справжні бактерії .
Архебактерії вважаються найпримітивнішою групою на нашій планеті. Всі вони є облігатними гетеротрофами, позбавлені пептидогліканових (муреїнових) клітинних покривів, які притаманні більшості евбактерій, нуклеїнові кислоти архебактерій частково метильовані, генетичний код не повністю ідентичне до інших організмів. Архебактерії здатні до фіксації молекулярного азоту, проте не здатні засвоювати складні органічні речовини, оскільки не мають гідролітичних екзоферментів. Тому серед архебактерій немає паразитичних та патогенних видів. Архебактерії населяють виключно екстремальні біотопи, що також непрямим чином підтверджує їх давність. До цього царства входить лише один відділ бактерій - Mendosicutes. Нижчих рослин серед архебактерій немає.
До царства справжніх бактерій - Eubacteria - належать всі інші відомі на планеті прокаріоти. Евбактерії здатні продукувати гідролітичні екзоферменти, і тому серед них відомо чимало паразитичних та патогенних представників. Генетичний код евбактерій вельми схожий з тим, який мають евкаріоти. Більшість евбактерій, за винятком мікоплазм, має клітинні стінки, до складу яких входить муреїн.
До Eubacteria відносять чотири відділи: Mollicutes - мікоплазми, Gracilicutes - грам-негативні бактерії, Firmacutes - грам-позитивні бактерії, та Cyanophyta (Cyanobacteria) - синьозелені водорості, або ціанобактерії. Останній відділ - Cyanophyta - представляє таксон, до якого входять види, що здійснюють оксигенний фотосинтез, і тому традиційно синьозелені водорості вважають нижчими рослинами і розглядають в курсі ботаніки.
Походження евкаріот
Існування прірви, яка відділяє прокаріот від евкаріот, не знаходила пояснень з точки зору еволюційних постулатів, сформульованих у ХІХ ст. Ч. Дарвіном. Згідно з теорією Дарвіна, різноманітність органічного світу пов'язана з поступовим еволюційним процесом, передумовою для якого було виникнення різноманітності пристосувальних ознак на основі випадкової спадкової мінливості, а рушійною силою - природній відбір з цієї різноманітності лише тих організмів, яким ці нові ознаки забезпечували переваги в конкурентній боротьбі над іншими близько спорідненими особинами. Тому з точки зору класичного дарвінізму, між типовими прокаріотами та типовими евкаріотами мали б існувати перехідні форми, яких проте навіть наприкінці ХХ ст. виявити не вдалось.
Автогенетична гіпотеза походження евкаріот. У ХІХ ст та першій половині ХХ ст. переважна більшість біологів обминала питання походження евкаріот, вважаючи, що вони виникли внаслідок дарвінівської еволюції, проте перехідні форми вимерли. Гіпотеза, що описувала походження евкаріот за таким сценарієм, отримала назву автогенетичної гіпотези (рис. 1.2).
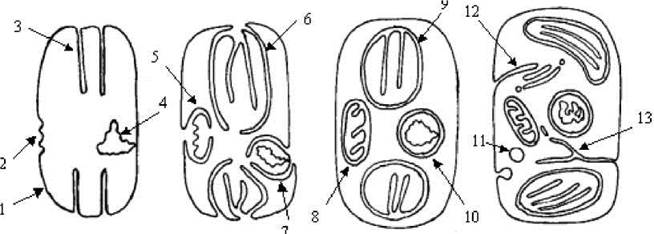
Рис. 1.2. Автогенетична гіпотеза походження евкаріот (зліва направо - послідовні етапи еволюції прокаріотичної клітини через розвиток системи інвагінацій плазмалеми, які врешті решт призводять до утворення евкаріотичної клітини. 1 - плазмалема; 2 - ділянка плазмалеми, що містить ферменти дихального ланцюгу; 3 - інвагінація плазмалеми, в яку вбудовані молекули хлорофілу - тилакоїд; 4 - нуклеоїд; 5 - інвагінація плазмалеми, що починає утворювати оболонку мітохондрії; 6 - інвагінація плазмалеми, що починає утворювати оболонку пластиди; 7 - інвагінація плазмалеми, що починає утворювати ядерну оболонку; 8 - мітохондрія; 9 - пластида; 10 - ядро; 11 - лізосома, що є похідною від травної вакуолі; 12 - комплекс Гольджі, що утворюється внаслідок інвагінації плазмалеми; 13 - ендоплазматична сітка, що утворюється внаслідок інвагінації плазмалеми.
Згідно з автогенетичною гіпотезою, евкаріотична клітина утворилась внаслідок розвитку плазмалемою системи інвагінацій, які надалі замкнулись навколо ділянок плазмалеми з ферментами дихального ланцюгу, фотосинтетичними пігментами, нуклеоїда, утворивши мітохондрії, пластиди та ядро, відповідно. Розвиток системи інвагінацій мімбран призвів також до виникнення ендоплазматичної сітки, комплексу Гольджі, лізосом та ін. Проте автогенетична гіпотеза не змогла пояснити чимало фактів. Наприклад, чому мітохондрії та хлоропласти містять власну ДНК і розмножуються поділом, чому в мітохондріях та хлоропластах наявні легкі прокаріотичні рибосоми, чому поровий апарат мітохондрій та пластид подібний до
порового апарату прокаріот, і відрізняється від порового апарату плазмалеми евкаріот, як виник мітоз та з'явились мікротрубочки, та ін.
Ендосимбіотична гіпотеза походження евкаріот. Наприкінці ХІХ ст. видатний ботанік та фізіолог рослин М. Цвет, на основі спостережень поділу хлоропласту та ядерця у зеленої водорості Spirogyra висловив припущення, що клітини рослин можливо представляють собою симбіотичний утвір з клітини-господаря та бактеріальних внутршньоклітинних симбіонтів, які відомі як хлоропласти та ядерця. Ця припущення на початку ХХ ст. було розвинене та опубліковане зоологом Фамінциним. У 20-40-х роках гіпотеза про ендосимбіотичне походження евкаріотичної клітини була детально розроблена К.С. Мережковським. Ці вчені на основі аналізу даних цитології та ліхенології продемонстрували, як симбіози могли надавати еволюційному процесу не поступового, а стрипкоподібного характеру та обумовити виникнення евкаріотичної клітини поминаючи перехідні форми між типово прокаріотичним та евкаріотичним планами будови. Проте роботи К.С. Мережковського не знайшли підтримки у широкого загалу біологів. Гіпотезу ендосимбіотичного походження евкаріот називали «вражаючим витвіром людської фантазії», «луна-парком», «непристойною темою у колі освідчених біологів».
У 60-х роках гіпотеза ендосимбіотичного походження евкаріотичної клітини була заново сформульована молодою американською дослідницею - Л. Маргеліс. Наводячи докази ендосимбіотичного походження мітохондрий та пластид Л. Маргеліс широко використала дані електронно-мікроскопічних досліджень, яких ще не міг мати в своєму розпорядженні К.С. Мережковський.
Гіпотеза ендосимбіотичного походження евкаріотичної клітини, яку спочатку називали гіпотезою Маргеліс, викликала чимало запеклих дискусій, і ініціювала цілу низку найрізноманітніших досліджень щодо її перевірки. Слід відмітити, що сама Л. Маргеліс, ознайомившись у 70-х роках з працями К.С. Мережковського, була настільки вражена співпадінням основних положень з її власними висновками, що запропонувала називати гіпотезу, названу на її честь, гіпотезою Мережковського. Сьогодні роль симбіозів в походженні мітохондрій та пластид вважається доведеною, а перший варіант гіпотези ендосимбіотичного походження евкаріотичної клітини отримав в біології назву гіпотези Мережковського-Маргеліс, або ортодоксальної ендосимбіотичної гіпотези.
Згідно з гіпотезою Мережковського-Маргеліс, евкаріотична клітина виникла внаслідок кількох ендосимбіозів (рис. 1.3). Гіпотетична прокаріотична анаеробна клітина, здатна до фагоцитозу, захопила, проте не перетравила, а зберігла в цитоплазмі аеробну гетеротрофну бактерію, яка надалі трансформувалась в мітохондрію. Далі клітина-господар, що містила мітохондрію, встурила в симбіоз з рухливою спірохетоподібною гетерофною бактерією, яка дала початок джгутику. Після цього внаслідок автогенетичного процесу, пов'язаного з утворенням глибоких інвагінацій плазмалеми, навколо нуклеоїду клітини-господаря утворилось ядро. Далі базальні тіла джгутиків трансформувались в центри організації веретена поділу, внаслідок чого виник спочатку мітоз, і його модифікація - мейоз разом із статевим процесом. Виникла перша гетеротрофна еукаріотична клітина.
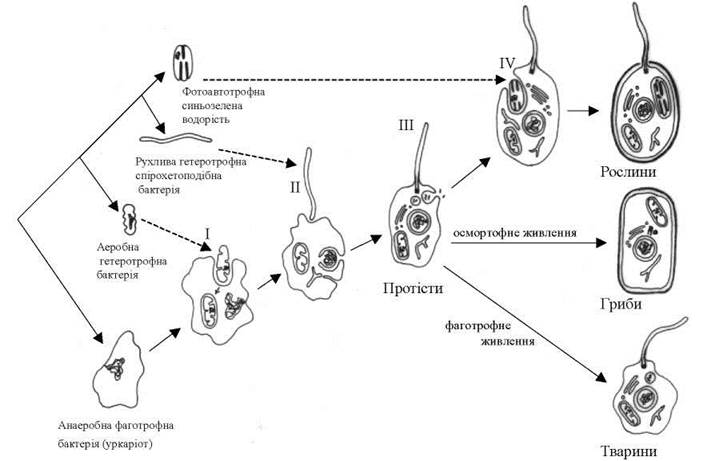
Рис. 1.3. Походження евкаріотичної клітини згідно з теорією Мережковського- Маргеліс. Пунктирною лінією позначено симбіотичні події (І - симбіоз, що призвів до утворення мітохондрії; ІІ - симбіоз, що призвів до утворення джгутика; ІІІ - автогенетичне утворення ядра та одномембранних евкаріотичних органел: ендоплазматичної сітки, комплексу Гольджі, лізосом; ІV - симбіоз, що призвів до утворення пластиди).
Нащадки цієї клітини, еволюція яких пішла по шляху вдосконалення фагоцитозу, дали початок тваринам. Ті ж нащадки, які еволюціонували в напрямку удосконалення осмотрофного живлення, дали початок грибам. А симбіоз гетеротрофної евкаріотичної клітини з синьозеленою водороістю призвів до появи рослин, оскільки симбіотична синьозелена водорість надалі трансформувалась у пластиду.
Численні перевірки ендосимбіотичної гіпотези, виконані з широким використанням електронної мікроскопії та мікрохімічних методів, з одного боку, не підтвердили ряд положень ортодоксальної ендосимбіотичної гіпотези - зокрема, положення про симбіотичне походження джгутику. З іншого боку, молекулярні дослідження, виконані у 90-роках суттєво змінили погляди на час виникнення ядра: вони підтвердили автогенетичну гіпотезу, і навіть більше - показали, що на довгому шляху, який врешті решт призвів до появи на Земній кулі евкаріот, однією з перших подій була саме поява морфологічно оформленої ядерної оболонки.
Таким чином, на початку ХХІ ст. теорія походження евкаріот сформувалась на основі синтезу обох гіпотез - як автогенетичної, так і ендосимбіотичної.
Синтетична гіпотеза походження евкаріот. У створенні сучасної системи органічного світу та розвитку уявлень про походження та напрямки еволюції евкаріот надзвичайно важливу роль відіграли молекулярно-систематичні дослідження різних груп водоростей та грибів, а також найпростіших, виконаних у 90-х роках ХХ ст. Методами молекулярної таксономії було показано, що найпримітивнішими евкаріотами є невелика група одноклітинних гетеротрофних джгутиконосців - діпломонади, оксимонади та ретортамонади, у яких немає мітохондрій, проте є морфологічно оформлене ядро, ендоплазматична сітка, джгутики, мітоз, мейоз та статевий процес. За результатами аналізу ряду генів (в першу чергу, того, що кодує малу субодиницю рибосомальної РНК), ці джгутиконосці виявились більш спорідненими з архебактеріями, ніж з евбактеріями.
Сьогодні вважають, що першим кроком на шляху виникнення евкаріот було утворення у гіпотетичного прокаріотичного пращура - уркаріоти - численних внутрішніх інвагінацій плазмалеми, які, з одного боку, замкнули прокаріотичний нуклеоїд у двомембранну оболонку (тобто утворили морфологічно оформлене ядро), а з іншого - призвели до утворення ендоплазматичної сітки та похідного від неї комплексу Гольджі, а також травних вакуолей та їх похідних - лізосом (рис. 1.4).
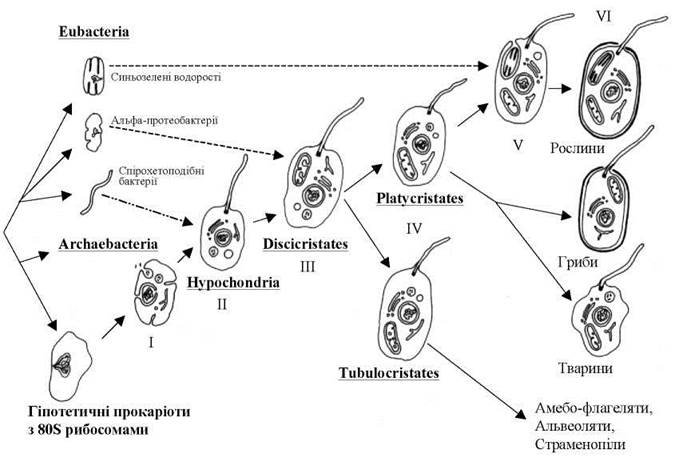
Рис. 1.4. Виникнення евкаріот та їх дивергенція згідно з синтетичною гіпотезою. Суцільними стрілками позначені напрямки еволюції клітини, штриховими стрілками - ендосимбіози, штрихпунктирною стрілкою - горизонтальний перенос гену, що кодує тубулін, внаслідок чого виникли мікротрубочкові клітинні системи. І - автогенетичне утворення ядра та одномембранних евкаріотичних органел; ІІ - виникнення мікротрубочкових систем, джгутикового апарату, мітозу, мейозу, статевого процесу; ІІІ - ендосимбіоз, внаслідок якого виникли мітохондрії з дископодібними кристами; IV - виникнення платикристат та тубулокристат; V - ендосимбіоз, внаслідок якого виникла первинна пластида; VI - велика радіація евкаріот та виникнення в межах платикристат рослин, грибів та тварин, а в межах тубулокристат - амебо-флагелят, альвеолят та страменопілів.
На другому етапі внаслідок горизонтального переносу гену, що кодує білок тубулін від спірохетоподібних бактерій, примітивні евкаріоти набули здатності до синтезу тубулінових мікротрубочок1. В результаті у евкаріот з'явився цитоскелет, джгутики з базальним тілами, веретено поділу, мітоз. Базальні тіла джгутиків надалі у частини представників трансформувалися у клітинний центр, а порушення нормального мітозу (зокрема, скорочення інтерфази) призвели до виникнення мейозу і пов'язаного з ним статевого процесу. Група первинно безмітохондріальних евкаріот сьогодні розглядається як перше, найпримітивніше царство евкаріот - Hypochondria (домітохондріальні, або гіпохондріати). Всі гіпохондріати є одноклітинними гетеротрофами з тваринною стратегією живлення.
На третьому етапі евкаріотична клітина утворила симбіотичний комплекс з прокаріотичною клітиною, схожою із сучасними альфа-протеобактеріями. Ця прокаріотична клітина надалі трансформувалась у мітохондрію. За результатами як цитологічних, так і молекулярних досліджень, найдавнішими мітохондріальними евкаріотами вважаються ті, які мають мітохондрії з особливими дископодібними кристами. Група мітохондріальних евкаріот з дископодібними кристами прийнята як таксон рангу царства - Discicristates (дискокристати). В основі царства дискокристат знаходяться первинно гетеротрофні організми.
На четвертому етапі евкаріоти розділилися на дві великих групи. Одна з цих груп має мітохондрії з трубчастими кристами, і складає царство Tubulocristates (тубулокристати, трубчастокристні), друга - мітохондрії з переважно пластинчастими кристами, і приймається як царство Platycrsitates (платикристати, пластинчастокристні). Подібно до дискокристат, в основі філ тубуло- та платикристат знаходяться первинно гетеротрофні організми із тваринною стратегією живлення.
На п'ятому етапі в евкаріотичному світі з'явилися перші рослини. За молекулярними та цитологічними даними, ця подія пов'язана з симбіозом гетеротрофної евкаріоти-платикристати з фотоавтотрофним прокаріотом - синьозеленою водорістю. Як наслідок цього симбіозу утворилась пластида, одягнена двома мембранами, яка отримала назву первинно симбіотичної пластиди. Подальша дивергенція організмів з первинно симбіотичними пластидами обумовила виникнення у межах філи платикристат групи фотоавтотрофних відділів, як склали подцарство Plantae - рослини. Дивергенція гетеротрофних платикристат за типами живлення обумовила виникнення з одного боку, відділів з осмотрофним живленням - підцарства Fungi (гриби), з іншого - таксонів з фаготрофним живленням, що склали підцарство Animalia - тварини.
Велика радіація тубулокристат відбувалась майже паралельно з платикристатами, проте була пов'язана не стільки з еволюцією за типами живлення, скільки з еволюцією клітинних покривів та джгутикового апарату. В основі тубулокристат залишились амебоїдні організми, які складають підцарство Ameboflagellates (амебо-флогеляти). Дві більш продвинуті філи представляють, з одного боку, таксони зі специфічними, альвеольованими покривами - Alveolates (альвеоляти) та з іншого - зі специфічними субмікроскопічними тричленними волосками на поверхні клітин або джгутиків (т.з. ретронемами) - Stramenopiles (страменопіли).
Первинні та вторинні ендосимбіози. Ендосимбіози, при яких в евкаріотичну клітину-господаря включався прокаріотичний симбіонт, що надалі трансформувався в клітинну органелу, називають первинними. На основі молекулярно-таксономічних досліджень наприкінці ХХ ст. було встановлено, що в світі евкаріот первинні симбіози відбулися лише двічі. При першому ендосимбіозі прокаріотична бактерія, схожа з альфа-протеобактеріями, вступила в симбіоз з фаготрофним джгутиконосцем з царства гіпохондріат і трансформувалась у мітохондрію з трьома типами крист - дископодібними, трубчастими та пластинчастими. Подальша морфологічна еволюція мітохондрії відбувалася поступово, майже паралельно з клітиною-господарем. Так, у тубулокристат дископодібні кристи були редуковані, а зберіглися трубчасті та інколи разом з ними - пластинчасті кристи. У платикристат були втрачені як дископодібні, так і трубчасті кристи, а зберіглися лише пластинчасті (за винятком відділу Haptophyta).
Другий первинний ендосимбіоз відбувся між прокаріотичним оксигенним фотоавтотрофом - синьозеленою водорістю, та гетеротрофним організмом з царства платикристат. Як наслідок, виникла перша евкаріотична клітина, здатна до оксигенного фотосинтезу - так звана "протоводорость", тобто утворились евкаріотичні рослини. У "протоводорості" пластида була вкрита двома мембранами, з яких внутрішня являла собою мембрану прокаріотичного ендосимбіонту - синьозеленої водорості, а зовнішня була похідною від плазмалеми клітини-господаря, і ймовірно представляла собою видозмінену мембрану травної вакуолі, в яку потрапив ендосимбіонт.
Згідно до молекулярних даних, "протоводорість" дала початок трьом паралельним гілкам рослин з первинно-симбіотичними пластидами - глаукоцистофітовим водоростям (Glaucocystophyta), червоним водоростям (Rhodophyta) та зеленим водоростям (Chlorophyta). Всі ці три відділи зберігають пластиди, що оточені лише двомембранною оболонку. Глаукоцистофітові водорості зберігають цікаву атавістичну ознаку - між зовнішньою та внутрішньою мембраною пластиди розташовується шар муреїну - речовини, характерної для клітинних оболонок більшості евкабактерій, і зокрема - синьозелених водоростей. Відділ глаукоцстофітових є сліпою гілкою еволюції рослин.
Пластиди червоних водоростей - родопласти - також зберігають деякі яскраві ознаки спорідненості із синьозеленими водоростями, зокрема - особливі так звані фікобілінові пігменти. Пластиди зелених водоростей - хлоропласти - зберігають найменше фенотипічних рис схожості із прокаріотичним ендосимбіонотом. Від зелених водоростей беруть початок вищі рослини, причому всі вони зберігають первинно-симбіотичні хлоропласти.
Червоні та зелені водорості неодноразово ставали ендосимбіонтами гетеротрофних евкаріот з різних царств - дискокристат, тубулокристат та платикристат. Симбіози, що відбувалися за сценарієм "евкаріотичний господар + евкаріотичний ендосимбіонт" називають вторинними.
Внаслідок вторинних ендосимбіозів виникла велика група відділів водоростей зі вторинно-симбіотичними пластидами. Вторинно симбіотичні пластиди, на відміну від первинно-симбіотичних, одягнені переважно чотирма мембранами, з яких внутрішня мембрана є залишком плазмалеми синьозеленої водорості (первинного ендосимбіонту), друга мембрана - видозміненою мембраною травної вакуолі господаря первинного ендосимбіонта (червоної або зеленої водорості), який є водночас вторинним ендосимбіонтом, третя мембрана - власне плазмалемою вторинного ендосимбіонта, четверта, зовнішня мембрана - плазмалемою господаря вторинного ендосимбіонта.
Цитоплазма, яка знаходиться у проміжку між другою та третьою мембраною, так званий перипластидний простір, є редукованою цитоплазмою вторинного ендосимбіонта, і у деяких випадках зберігає 80S рибосоми та редуковану ендоплазматичну сітку. Зовнішні (третю та четверту) мембрани вторинно- симбіотичних пластид називають хлоропластною ендоплазматичною сіткою, і позначають абревіатурою CER (від англ. Chloroplast endoplasmatic reticulum).
Проте особливо цікавим є той факт, що у двох відділах водоростей зі вторинно- симбіотичними пластидами - хлорарахнієфітових (Chlorarachniophyta) та криптофітових (Cryptophyta) - у перипластидному просторі зберігається навіть редуковане ядро вторинного ендосимбіонта - нуклеоморф. Нуклеоморф оточений двомембранною оболонкою і місить ДНК, яка по наявним у нуклеоморфі генам у хлорарахнієфітових споріднена з ядерною ДНК зелених водоростей, а у криптофітових - з ядерною ДНК червоних водоростей.
У деяких водоростей, зокрема - у евгленофітових - вторинно симбіотичні пластиди вкриті не чотирма, а лише трьома мембранами. Вважається, що у цих водоростей одна з зовнішніх мембран пластиди була вторинно втрачена.
Відомі випадки, коли пластиди (як первинно-, так і вторинно-симбіотичні) редукувалися, і тоді виникали вторинно гетеротрофні таксони. Зокрема, доведена вторинна редукція пластиди у тварин з типу Apicomplexa, пращурами яких були фотоавтотрофні динофітові водорості (Dinophyta).
Є також дані, які дозволяють припустити, що оомікотові гриби (Oomycota) утворились внаслідок вторинного переходу до гетеротрофного типу живлення від однієї з груп водоростей-тубулокристат.
Крім того, вторинно гетеротрофні таксони низьких та середніх рангів (від видів до порядків і навіть класів) відомі майже в кожному відділі водоростей, а також серед вищих рослин.
Поширення пластид через вторинні ендосимбіози обумовило вининкнення значної кількості відділів водоростей. Наприклад, якщо водорості з первинно симбіотичними пластидами представлені лише трьома відділами (Glaucocystophyta, Rhodophyta, Chlorophyta), то водоростей зі вторинно симбіотичними пластидами належать до 12 відділів. Серед них два відділи зберігають нуклеоморф (Chlorarachniophyta, Cryptophyta), 8 відділів мають чотирьохмембранні вторинно симбіотичні пластиди без нуклеоморфу, що походять від червоних водоростей (Raphidophyta, Chrysophyta, Eustigmatophyta, Xanthophyta, Phaeophyta, Bacillariophyta, Dictyochophyta, Haptophyta). Евгленофітові водорості мають тримембранні пластиди, пращурами яких є зелені водорості. У Dinophytaпластиди утворювались неодноразово, як внаслідок симбіозів з зеленими водоростями, так і навіть внаслідок третинних симбіозів зі вторинно симбіотичними фотоавтотрофами.
Склад водоростей на рівні відділів
Внаслідок процесів та подій, розглянутих у розділах 2-3, організми, відомі під традиційною назвою "водорості", потрапили у найрізноманітніші філи органічного світу. Сьогодні біологи виділяють 16 відділів водоростей. До водоростей з філогенетичної точки зору слід віднести також один тип тварин (Apicomplexa), який вторинно походить від водоростей внаслідок редукції пластиди, проте зберігає пластидний геном.
Згідно зі схемою, що відображує напрямки еволюції клітин-господарів (рис. 1.5), водорості належать до двох надцарств: Procaryota (1 відділ) та Eucaryota (14 відділів).
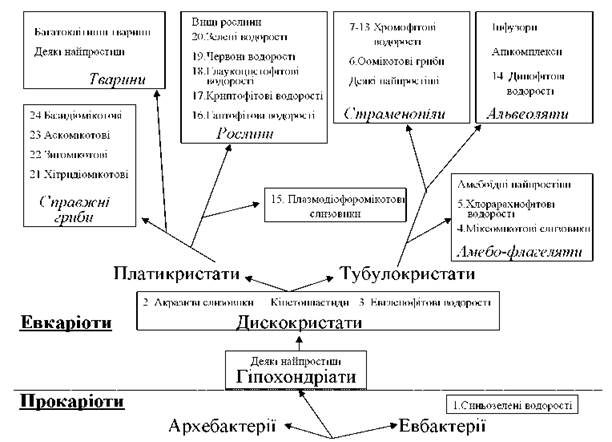
Рис. 1.5. Місце водоростей у системі органічного світу. Номерами позначені відділи та групи відділів, які прийнято вважати водоростями.
Серед нижчих рослин 16 відділів включають фотоавтотрофних представників, тобто представляють водорості. З них один відділ - Cyanophyta - належить до прокаріот, три відділи - Glaucocystophyta, Rhodophyta, Chlorophyta - до водоростей з первинно симбіотичними пластидами, решту представляють відділи зі вторинно симбіотичними пластидами.
Нижче подано систематичне положення відділів нижчих рослин у системі органічного світу та стисла характеристика кожного з відділів.
Надцарство Прокаріоти - Procaryota
Царство Еубактерії - Eubacteria
1. Cyanophyta - синьозелені водорості
Прокаріотичні фотоавтотрофні рослини
Надцарство Евкаріоти - Eucaryota
Царство Дискокристати - Discicristates
Представники царства характеризуються наявністю різноманітних крист, серед яких обов'язково представлені кристи дископодібної форми. Поділ ядра представлений закритим ортомітозом.
2. Euglenophyta - евгленофітові водорості
Евкаріотичні первинно гетеротрофні, фотоавтотрофні та вторинно гетеротрофні дискокристати, що мають клітинний покрив, представлений пелікулою. Фотоавтотрофні представники мають вторинно симбіотичні пластиди хлорофітного типу.
Царство Тубулокристати - Tubulocristates
Представники царства характеризуються наявністю трубчастих, перешнурованих при основі мітохондріальних крист. Всі фотоавтотрофні представники мають пластиди виключно вторинно-симбіотичного типу.
Підцарство Амебо-флагелляти - Ameboflagellatae
Об'єднує організми, у яких клітини голі, і в життєвому циклі представлені лише амебоїдні або монадні стадії (причому амебоїдна стадія, як правило, переважає). Джгутикові стадії позбавлені тричленних мастигонем - ретронем.
3. Chlorarachniophyta - хлорарахніофітові водорості.
Евкаріотичні фотоавтотрофні тубулокристати, у яких клітини голі, а вегетативне тіло представлене амебоїдами, здатними об'єднуватися у плазмодії. Пластиди вторинно симбіотичні, хлорофітного типу, з нуклеоморфом.
Підцарство Страменопіли - Stramenopiles
Об'єднує організми з різноманітними типами покривів (оболонками, панцирами, плазмалемою з додатковими захисними структурами). Головна ознака - наявність у клітини (переважно - на поверхні джгутиків) особливих тричленних мастигонем - ретронем, утворення яких починається або між мембранами ядерної оболонки, або між мембранами хлоропластної ендоплазматичної сітки.
Група відділів Хромофітові водорості.
Об'єднує всіх фотоавтотрофних страменопілів. Група монофілітична за всіма геномами - ядерним, мітохондріальним, пластидним. Для всіх відділів спільною ознакою є наявність вторинно симбіотичних пластид родофітного типу. Представники всіх відділів хромофітових водоростей мають хлорофіли а та с, чотиримембранні хлоропласти, в яких дві зовнішні мембрани утворюють хлорпластну ендоплазматичну сітку, ретронеми утворюються як між мембранами ядерної оболонки, так і між мембранами хлоропластної ендоплазматичної сітки. Відміни між відділами пов'язані, в першу чергу, з будовою клітинних покривів, наявністю фукоксантину, продуктами асиміляції, будовою фоторецепторного апарату, особливостями організації цитоскелету, а також з типами морфологічної структури тіла.
4. Rhaphidophyta - рафідофітові водорості.
Евкаріотичні фотоавтотрофні тубулокристати, у яких клітини голі (проте не амебоїдні). Пластиди вторинно симбіотичні, родофітного типу. Джгутикові стадії мають ретронеми. Продукт асиміляції - олія. Характерні особливості - наявність глотки та супрануклеарного апарату, представленого особливою видозміною комплексу Гольджі, а також відсутність стигми та структурного зв'язку між ядерною оболонкою та хлоропластною ендоплазматичною сіткою. Всі представники мають виключно монадний тип структури тіла.
5. Chrysophyta - золотисті водорості
Евкаріотичні фотоавтотрофні та вторинно гетеротрофні тубулокристати, у яких клітини голі (з кремнеземовими лусочками або без них) або вкриті пектиновою оболонкою. Пластиди вторинно симбіотичні, родофітного типу. Джгутикові стадії мають ретронеми. Продукт асиміляції - хризоламінарин. Характерна особливість - наявність у життєвому циклі стадії ендогенних кремнеземових цист.
6. Eustigmatophyta - евстигматофітові водорості
Евкаріотичні фотоавтотрофні тубулокристати, у яких клітини вкриті пектиновою оболонкою. Пластиди вторинно симбіотичні, родофітного типу. Джгутикові стадії мають ретронеми. Продукт асиміляції - хризоламінарин. Характерна особливість - наявність у монадних стадій унікального фоторецепторного апарату, розташованого у цитоплазмі біля основи джгутиків. Всі представники мають виключно кокоїдний тип структури тіла.
7. Xanthophyta - жовтозелені водорості
Евкаріотичні фотоавтотрофні тубулокристати, у яких клітини вкриті пектиновою або целюлозно-пектиновою оболонкою. Пластиди вторинно симбіотичні, родофітного типу. Джгутикові стадії мають ретронеми. Продукт асиміляції - хризоламінарин. Характерна особливість - відсутність жовтого ксантофілу фукоксантину.
8. Phaeophyta - бурі водорості
Евкаріотичні фотоавтотрофні тубулокристати, у яких клітини вкриті целюлозно- пектиновою оболонкою, до складу якої входять альгінати. Пластиди вторинно симбіотичні, родофітного типу. Джгутикові стадії мають ретронеми. Продукт асиміляції - ламінарин. Характерна особливість - виключно багатоклітинна будова таломів.
9. Bacillariophyta - діатомові водорості
Евкаріотичні фотоавтотрофні тубулокристати, у яких клітини вкриті кремнеземовим панциром. Пластиди вторинно симбіотичні, родофітного типу. Монадні стадії нерухомі, джгутики мають ретронеми. Продукт асиміляції - хризоламінарин. Характерні особливості - наявність кремнеземового панциру та виключно диплофазний життєвий цикл з особливою стадією ауксоспор.
10. Dictyochophyta - диктохофітові водорості
Евкаріотичні фотоавтотрофні та вторинно гетеротрофні тубулокристати, у яких клітини голі (у частини представників мають внутрішній кремнеземовий скелет). Пластиди вторинно симбіотичні, родофітного типу. Монадні стадії мають ретронеми. Продукт асиміляції - хризоламінарин. Характерна особливість - асоційованість базальних тіл джгутиків безпосередньо з ядерною мембраною, без участі джгутикових коренів.
Підцарство Альвеоляти - Alveolatae
Об'єднує організми, у яких клітини мають особливі - альвеольовані - покриви, в утворенні яких значну роль відіграють мікротрубочки. Для альвеолят характерні різні варіанти ядерного апарату, які помітно відрізняються від типової схеми будови ядра евкаріот. Ретронеми у альвеолят відсутні.
11. Dinophyta - динофітові водорості
Евкаріотичні первинно гетеротрофні, фотоавтотрофні та вторинно гетеротрофні тубулокристати, я яких клітини вкриті особливим типом покривів - альвеольованою амфієсмою. Пластиди вторинно симбіотичні і вельми різноманітні - хлорофітного та родофітного типів. У примітивних представників ядро типово евкаріотичне, у еволюційно продвинутих представляє особливий варіант ядерного апарату - динокаріон.
Додаток: тип Apicomplexa - апікомплекси
Евкаріотичні вторинно гетеротрофні облігатні паразити, які втратили мітохондрії, проте зберігають редуковані пластиди та пластидний геном.
Царство Платикристати - Platycristates
Представники царства характеризуються наявністю пластинчастих (інколи - також трубчастих) неперешнурованих при основі мітохондріальних крист.
Підцарство Рослини - Plantae
12. Haptophyta - гаптофітові водорості
Евкаріотичні фотоавтотрофні та вторинно гетеротрофні платикристати, що мають мітохондрії з трубчастими кристами (проте не перетягнутими при основі), клітинний покрив, представлений плазмалемою, що з зовнішнього боку вкрита субмікроскопічними органомінеральними лусочками, а з внутрішнього підстелена оперезуючою цистерною ендоплазматичної сітки. Пластиди вторинно симбіотичні, родофітного типу.
13. Cryptophyta - криптофітові водорості
Евкаріотичні фотоавтотрофні та вторинно гетеротрофні платикристати, що мають клітинний покрив, представлений перипластом. Пластиди вторинно симбіотичні, родофітного типу, з нуклеоморфом.
14. Glaucocystophyta - глаукоцистофітові водорості
Евкаріотичні фотоавтотрофні платикристати, що мають амфієсмоподібний клітинний покрив. Пластиди первинно симбіотичні, глаукоцистофітного типу (цианели), між зовнішньою та внутрішньою мембранами хлоропластної оболонки зберігається шар муреїну. Продукт асиміляції - крохмаль, що відкладається у цитоплазмі.
15. Rhodophyta - червоні водорості
Евкаріотичні фотоавтотрофні платикристати, клітини яких вкриті целюлозно- пектиновою оболонкою з фікоколоїдами. Пластиди первинно симбіотичні, родофітного типу (родопласти), з поодинокими тилакодами. Продукт асиміляції - багрянковий крохмаль, що відкладається у цитоплазмі. Специфічна ознака - повна відсутність джгутикових стадій.
16. Chlorophyta - зелені водорості
Евкаріотичні фотоавтотрофні платикристати, вкриті плазмалемою або клітинною оболонкою. Пластиди первинно симбіотичні, хлорофітного типу (хлоропласти). Продукт асиміляції - крохмаль, що відкладається у хлоропласті.
1 Сьогодні у біологів немає одностайної думки стосовно того, який комплекс органел виник раніш - ядерний чи мікротрубочковий. По цьому питанню існують дві альтернативні гіпотези - про первинність ядра і вторинність мікротрубочкової системи, і навпаки.