МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 5. ВЕГЕТАТИВНЫЕ ОРГАНЫ РАСТЕНИЙ
5.3. Стебель как вегетативный орган
Стебель — это осевой вегетативный орган, который обладает (в типичном случае) отрицательным геотропизмом и несет листья, почки и цветки. Стебель, как и корень, выполняет механическую (поддерживает листья) и проводящую функции. У травянистых растений зеленые стебли способны к фотосинтезу. Эти функции являются основными. Кроме основных функций стебель под влиянием условий существования может метаморфизироваться и выполнять дополнительные функции — запасающую, функцию размножения и др.
5.3.1. Первичное анатомическое строение стебля
Формирование анатомической и морфологической структуры стебля происходит благодаря верхушечной меристеме конуса нарастания стебля. Конус нарастания стебля отличается от конуса нарастания корня: нет корневого чехлика, он защищен зачаточными листьями, есть отличия в дифференцировке верхушечной меристемы: она дифференцирована не на три зоны, а на две — тунику и корпус (согласно теории, Дж. Будера и А. Шмидта) или мантию и корпус (по теории А. Фостера) (см. рис. 13).
Туника довольно тонкая, клетки самого наружного ее слоя делятся тангентально и образуют впоследствии эпидермис, т. е. этот слой меристемы фактически является протодермой и может рассматриваться как аналог дерматогена корня. Остальные клетки туники делятся в разных направлениях и образуют первичную кору стебля. Из меристемы корпуса формируется центральный цилиндр.
Таким образом, молодой стебель, как и молодой корень, дифференцирован на две топографические зоны: первичную кору и центральный цилиндр. Однако формирование первичного анатомического строения стебля и его строение на ранних этапах онтогенеза не только отличается от первичного анатомического строения корня, но имеет в пределах различных таксонов покрытосеменных растений специфические особенности. В частности, у многих однодольных растений (злаки) в отличие от двудольных первичная кора стебля выражена слабо и нечетко отделена от центрального цилиндра. У двудольных образуется эустель, у однодольных — атактостель. У однодольных при формировании сосудисто-волокнистых пучков молодого стебля весь прокамбий расходуется на образование первичной ксилемы и первичной флоэмы. У двудольных растений между первичными ксилемой и флоэмой сохраняется прокамбий, который впоследствии превращается в камбий.
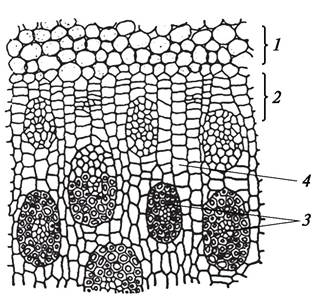
Эпидермис молодого стебля имеет характерные для этой ткани черты. У двудольных под эпидермисом находится первичная кора (рис. 84). Периферическая часть первичной коры стебля двудольных растений представлена колленхимой. У вегетативных побегов это чаще всего пластинчатая колленхима (подсолнечник однолетний — Helianthus annuus), у цветоносных (чеснок — Allium sativum) рыхлая. Клетки колленхимы содержат хлоропласты, и, следовательно, кроме арматурной функции у молодых растений колленхима выполняет функцию фотосинтеза. У однодольных растений под эпидермисом развивается склеренхима (рожь посевная — Secale cereale), реже уголковая колленхима (зебрина повислая — Zebrina pendula).
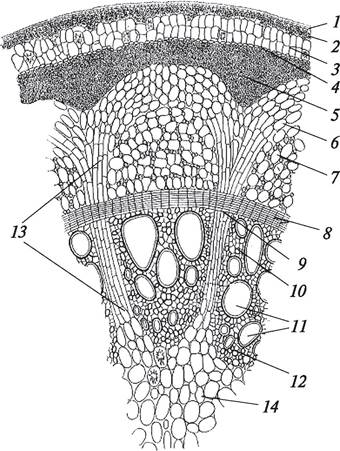
Рис. 84. Первичное анатомическое строение стебля подсолнечника (Helianthus annuus). Начальный этап перехода к вторичному строению: 1—сердцевина; 2—первичная кора; 3—эпидермис; 4—коллатеральный сосудисто-волокнистый пучок с развивающимся пучковым камбием; 5—формирующийся межпучковый камбий
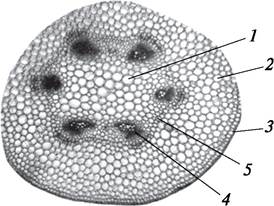
Подавляющая часть первичной коры двудольных растений образована паренхимой разных типов. За колленхимой располагается хлорофиллоносная паренхима (хлоренхима), ближе к центральному цилиндру — запасающая. Самый внутренний слой первичной коры состоит из клеток, заполненных крахмалом, и называется крахмалоносным влагалищем. Это образование аналогично эндодерме корня. Крахмал крахмалоносного влагалища не расходуется даже тогда, когда растение «голодает», поэтому его называют оберегаемым. Крахмалоносное влагалище является, как и эндодерма корня, физиологическим барьером и регулирует передвижение воды и пластических веществ в горизонтальном направлении. В стеблях некоторых двудольных (сем. Бобовые — Fabaceae) и однодольных (сциндапсус пятнистый — Scindapsuspictus) образуется типичная эндодерма.
Крахмалоносное влагалище граничит с перициклом центрального цилиндра, который возникает из периферического слоя корпуса благодаря тангентальным делениям меристемы и образован одним слоем клеток таблитчатой (паренхиматозной) формы. Это специфическая первичная меристема, которая ограничивает центральный цилиндр стебля. Центральная часть клеток корпуса (часто называемая основной меристемой) дает начало паренхиме центрального цилиндра и васкулярной меристеме — прокамбию.
Прокамбий дифференцируется под перициклом или в виде обособленных групп (пучков), разграниченных паренхимой, или в виде сплошного кольца (рис. 85). Периферические участки прокамбия образуют первичную флоэму, т. е. она возникает экзархно, развивается центростремительно. Таким образом, протофлоэма формируется вблизи перицикла, метафлоэма располагается за ней ближе к центру стебля.
Рис. 85. Схема развития первичного анатомического строения стебля двудольных растений: 1—первичная кора; 2—центральный цилиндр; 3—перицикл; 4— прокамбиальные тяжи и прокамбий между первичной ксилемой и первичной флоэмой; 5—появление протофлоэмы; 6—появление протоксилемы; 7—сформированные первичные коллатеральные сосудисто-волокнистые пучки; 8—метафлоэма; 9—метаксилема. Стрелками показано направление развития элементов первичной флоэмы и первичной ксилемы
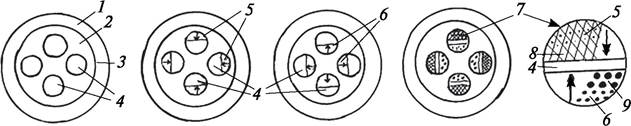
В этих же тяжах прокамбия или прокамбиальных кольцах образуется первичная ксилема, но она возникает эндархно и развивается центробежно. Таким образом, в молодых стеблях двудольных и однодольных растений образуются коллатеральные сосудисто-волокнистые пучки. У некоторых двудольных (лен посевной — Linum usitatissimum) возникают сплошные, примыкающие друг к другу кольца первичных флоэмы и ксилемы. В центре стели у двудольных образуется сердцевина из паренхимных клеток. У однодольных растений сердцевины нет, сосудисто-волокнистые пучки разбросаны диффузно по всему центральному цилиндру.
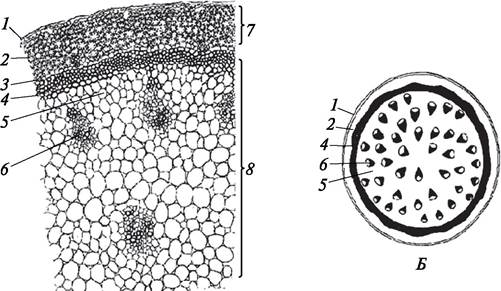
У травянистых однодольных растений первичная анатомическая структура стебля сохраняется в течение всей жизни (рис. 86).
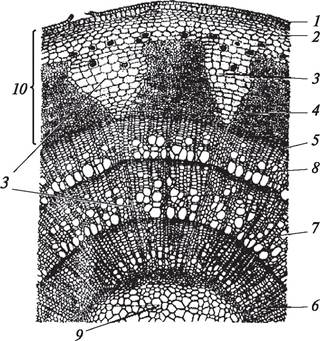
Рис. 86. Анатомическое строение (А) и схема (Б) поперечного разреза стебля ириса (Iris germanica): 1—эпидермис; 2—хлоренхима; 3—эндодерма; 4—перициклическая склеренхима; 5—основная паренхима; 6—закрытый коллатеральный пучок; 7—первичная кора; 8—центральный цилиндр (атактостель)
5.3.2. Переход от первичного анатомического строения стебля к вторичному. Вторичное анатомическое строение стеблей травянистых растений
У двудольных растений переход от первичной анатомической структуры стебля к вторичной связан с переходом прокамбия в пучковый камбий и появлением и деятельностью межпучкового камбия. Межпучковый камбий возникает на уровне пучкового камбия в результате дедифференцировки клеток паренхимы, разделяющей пучки.
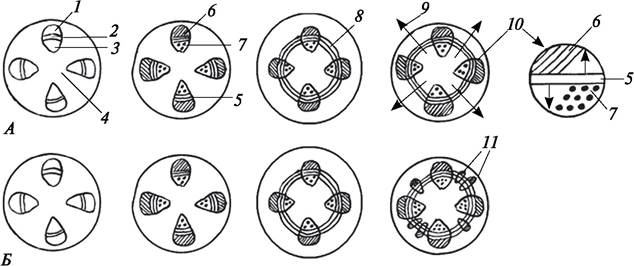
У травянистых двудольных растений выделяют три типа вторичного анатомического строения стебля — пучковый, промежуточный и непучковый (рис. 87).
Рис. 87. Схема развития вторичного анатомического строения стеблей двудольных растений. Изменения, происходящие в центральном цилиндре: А—пучковый тип строения; Б—промежуточный тип строения: 1— первичная флоэма; 2—прокамбий; 3—первичная ксилема; 4—сердцевина; 5—образовавшийся пучковый камбий и вторичные проводящие ткани; 6—вторичная флоэма; 7—вторичная ксилема; 8—появление межпучкового камбия; 9—образование первичных сердцевинных лучей; 10— сформированные коллатеральные сосудисто-волокнистые пучки; 11—добавочные сосудисто-волокнистые пучки, образованные межпучковым камбием
При развитии стебля пучкового типа пучковый камбий образует вторичную ксилему и флоэму, а межпучковый камбий производит только паренхиму, которая образует первичные сердцевинные лучи и соединяет сердцевину с периферическими участками стебля (кирказон крупнолистный — Aristolochia macrophylla) (рис. 88).
Рис. 88. Вторичное анатомическое строение стебля кирказона (Aristolochia clematitis) (стебель пучкового типа): 1—эпидермис; 2—колленхима; 3—паренхима первичной коры; 4—крахмалоносное влагалище; 5—склеренхима перициклического происхождения; 6—паренхима сердцевинного луча; 7—вторичная флоэма; 8—пучковый камбий;
9—межпучковый камбий; 10—вторичная ксилема; 11—сосуды вторичной ксилемы; 12—остатки первичной ксилемы; 13—первичный сердцевинный луч; 14—сердцевина стебля
При образовании стебля промежуточного типа четко выраженных сердцевинных лучей нет. Межпучковый камбий, кроме паренхимы, образует дополнительные коллатеральные пучки, которые располагаются между основными пучками и отличаются от них размерами. Основные пучки располагаются на более-менее определенном расстоянии один от другого, а расположение дополнительных пучков не носит какой -либо закономерности. В межпучковых зонах число их и размеры варьируют (подсолнечник однолетний — Helianthus annuus,полынь обыкновенная — Artemisia vulgaris) (рис. 89).
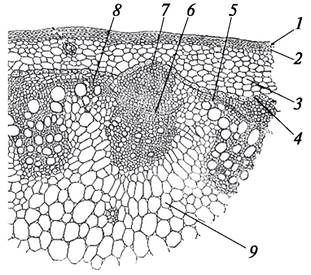
Рис. 89. Вторичное анатомическое строение стебля подсолнечника (Helianthus annuus) (стебель промежуточного типа): 1—эпидермис; 2—пластинчатая колленхима; 3— паренхима первичной коры; 4—крахмалоносное влагалище (эндодерма); 5—межпучковый камбий; 6—пучковый камбий; 7—основной проводящий пучок; 8—добавочный проводящий пучок; 9—паренхима сердцевины стебля
При переходе от первичного анатомического строения к вторичному стебель непучкового типа образуется в тех случаях, когда в стебле, имеющем первичное строение, прокамбий закладывался в виде сплошного кольца. В таких стеблях из прокамбия образуется сплошное кольцо камбия и, как следствие этого, сплошное кольцо вторичной флоэмы и вторичной ксилемы. От сердцевины к периферии стебля идут однорядные тяжи паренхимы, которые выполняют ту же функцию, что и сердцевинные лучи (лен посевной — Linum usitatissimum,вьюнок полевой — Convolvulus arvensis) (рис. 90).
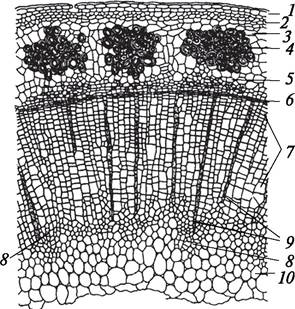
Рис. 90. Вторичное анатомическое строение стебля льна (Linum usitatissimum) (стебель непучкового типа): 1—эпидермис; 2—паренхима первичной коры; 3— крахмалоносное влагалище (эндодерма); 4—перициклическая склеренхима (лубяные волокна); 5—вторичная флоэма; 6—камбий; 7—вторичная ксилема; 8—остатки первичной ксилемы; 9—сердцевинные лучи; 10—сердцевина стебля
При переходе стеблей к вторичному анатомическому строению перицикл часто образует механическую ткань — перициклическую склеренхиму. Первичная кора претерпевает незначительные изменения.
5.3.3. Вторичное анатомическое строение стеблей древесных растений
Анатомическое строение стеблей древесных растений из класса Двудольные сохраняет план строения, подобный строению травянистых растений, однако есть ряд специфических особенностей (рис. 91). Так как камбий ежегодно активно функционирует, во вторичной ксилеме древесных растений образуются годичные кольца (кольца прироста). Вторичная ксилема и вторичная флоэма богаты механическими волокнами. В ксилеме это либриформ, во флоэме — лубяные волокна. В ксилеме в конце вегетационного периода образуется много склеренхиматозной паренхимы. У древесных растений хорошо выражены первичные сердцевинные лучи, образованные паренхимными клетками, соединяющие сердцевину стебля с первичной корой. Сердцевинные лучи могут состоять из лежачих клеток, вытянутых в радиальном направлении, и стоячих клеток, расположенных параллельно продольной оси стебля. Между клетками имеются узкие межклетники. В зависимости от типов клеток, которые входят в сердцевинный луч, различают первичные сердцевинные лучи гомоцеллюляр- ные (гомогенные) и гетероцеллюлярные (гетерогенные). Гомоцеллюлярные лучи состоят из одинаковых по морфологическим и функциональным особенностям лежачих клеток.
Рис. 91. Анатомическое строение трехлетней ветки липы (Tilia cordata): 1—перидерма с остатками эпидермиса; 2—паренхима первичной коры; 3—первичный сердцевинный луч; 4—вторичная флоэма; 5—камбий; 6, 7, 8—первый, второй и третий годичные приросты; 9—сердцевина стебля; 10—вторичная кора
Гетероцеллюлярные лучи образованы клетками двух типов — лежачими и стоячими. Первичные сердцевинные лучи бывают двурядными и многорядными. Они идут от сердцевины стебля до первичной коры. По сердцевинным лучам к центру стебля по радиусу переносится вода с раствором минеральных солей. По клеткам сердцевинных лучей происходит также передвижение пластических веществ. В различные сезонные периоды года пластические вещества по сердцевинным лучам передвигаются по-разному. Осенью они перемещаются в горизонтальном направлении от периферии к центру и откладываются в запас, а весной движутся из центра к периферии и переходят в ситовидные трубки флоэмы. По узким межклетникам первичных сердцевинных лучей осуществляется процесс газообмена.
Кроме первичных сердцевинных лучей, у древесных растений образуются вторичные сердцевинные лучи, которые, в отличие от первичных, начинаются слепо в каком-нибудь годичном кольце вторичной ксилемы и заканчиваются произвольно в любом участке вторичной ксилемы. Иногда они могут переходить во вторичную флоэму и заканчиваться в ней. Вторичные сердцевинные лучи всегда состоят из одного ряда клеток.
По мере старения растения претерпевает изменения и сердцевина (лат. medulla) стебля. Она дифференцируется на более рыхлую центральную часть, состоящую из расположенных продольными рядами паренхимных клеток, и перимедуллярную зону, которая образована мелкими, толстостенными, плотно прилегающими друг к другу клетками. У некоторых растений в сердцевине развиваются схизогенные вместилища.
Существенные изменения происходят с возрастом растения и в первичной коре древесных растений. На смену эпидермису формируется сначала перидерма, а затем на пятом — седьмом году жизни (у некоторых растений в более позднем возрасте) образуется ритидом. Иногда, характеризуя древесные растения, выделяют вторичную кору, под которой следует понимать всю совокупность тканей, расположенных к периферии от камбия (вторичная флоэма, остатки паренхимы первичной коры с включениями механических, секреторных тканей, перидерма).
У древесных однодольных растений (пальмы, юкки) утолщение стебля происходит атипично благодаря возникновению дополнительных меристематических зон, которые закладываются между первичной корой и центральным цилиндром (атактостелой) (рис. 92). Они функционируют в течение всей жизни растений. Эти меристематические зоны формируют в центростремительном направлении закрытые коллатеральные или концентрические амфивазальные пучки и паренхиму, т. е. образуют дополнительные участки паренхимы и элементы проводящей системы.
Рис. 92. Вторичное анатомическое строение стебля драцены окаймленной (Dracaena marginata): 1—первичная кора; 2—камбий и формирующийся вторичный прирост; 3 —сформировавшиеся вторичные амфивазальные сосудисто-волокнистые пучки; 4—паренхима вторичного прироста
5.3.4. Морфологические особенности стеблей
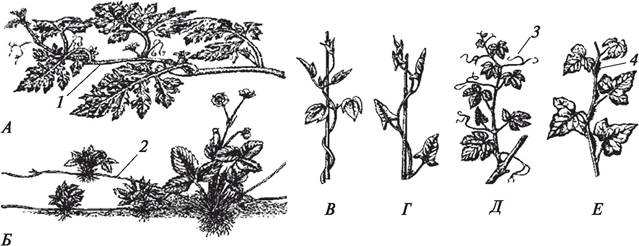
Морфологическое строение стеблей покрытосеменных растений характеризуется значительно большим разнообразием, чем анатомическое. По характеру и направлению роста можно выделить стебли прямостоячие, приподнимающиеся, вьющиеся, цепляющиеся, ползучие, стелющиеся, укороченные (рис. 93).
Рис. 93. Виды стеблей по характеру и направлению роста: А—стелющийся (арбуз обыкновенный — Citrullus vulgaris); Б—ползучий (земляника лесная — Fragaria vesca); В —вьющийся по часовой стрелке (хмель обыкновенный — Humulus lupulus); Г—вьющийся против часовой стрелки (вьюнок полевой — Convolvulus arvensis); Д—лазящий при помощи усиков (виноград культурный — Vitis vinifera); Е—лазящий при помощи корней- присосок (корней-прицепок) (плющ обыкновенный — Hedera helix): 1—плети; 2 —усы; 3—усики; 4—корни-присоски (корни-прицепки)
Прямостоячие стебли присущи подавляющему большинству растений. Они имеют достаточно хорошо развитые механические ткани, что позволяет им самостоятельно поддерживать вертикальное отрицательно геотропичное положение.
У растений с приподнимающимся стеблем (клевер луговой — Trifolium pratense) основание дуговидно изогнуто, а верхушка стебля располагается вертикально.
Вьющиеся стебли отличаются способностью к круговым (нутационным) движениям, что позволяет растениям обвиваться вокруг различных опор и принимать вертикальное положение. Круговые движения связаны с перемещением вокруг оси органа зон усиленного роста. Вьющиеся растения очень богаты гибереллином, и можно считать, что этот фитогормон причастен к осуществлению обвивания. Есть растения с правовращающимися (по часовой стрелке — жимолость-каприфоль — Lonicera caprifolium; хмель обыкновенный — Humulus lupulus) и левовращающимися (против часовой стрелки — ипомея пурпуровая — Ipomea purpurea, фасоль огненно-красная — Phaseolus coccineus, вьюнок полевой — Convolvulus arvensis) стеблями. В большинстве случаев растения обладают способностью к левовращению. Правое вращение стеблей отмечено у немногих видов.
При помощи различных приспособлений поддерживают стебель в вертикальном положении цепляющиеся растения. У винограда амурского (Vitis amurensis), винограда девичьего пятилисточкового (Partenocyssus quinquefolia) и других представителей семейства Виноградные стебель цепляется за опору при помощи усиков. Так же цепляются за опору и многие представители семейства Бобовые (горошек тонколистный — Vicia tenuifolia, чина клубненосная — Lathyrus tuberosus). У плюща обыкновенного (Hedera helix) на концах придаточных корней, которые в большом количестве образуются на стебле, возникают особые присоски. С помощью таких корней-присосок плющ прикрепляется к стволам деревьев, а в южных районах, где его используют для вертикального озеленения, — к стенам зданий.
При помощи черешков листьев, которые обвиваются вокруг опоры, поднимаются на высоту двух — четырех метров стебли многих видов ломоноса (Clematis).
Прямостоячие, приподнимающиеся, вьющиеся и цепляющиеся стебли являются ортотропными, они в конечном счете принимают вертикальное положение. Однако есть и плагиотропные стебли, располагающиеся горизонтально относительно поверхности почвы. К ним относятся ползучие и стелющиеся стебли.
Ползучими называются стебли, у которых в узлах образуются придаточные корни, с их помощью растение укореняется в субстрате и захватывает территорию — «ползет». Такие стебли характерны для лютика ползучего (Ranuncu lus repens), живучки ползучей (Ajuga reptans), земляники лесной (Fragaria vesca). Иногда ползучие стебли называют усами или надземными столонами.
В отличие от ползучих стелющиеся стебли не образуют придаточных корней и их олиственные или безлистные стебли распростерты по субстрату (грыжник голый — Herniaria glabra).Стелющиеся стебли встречаются как среди травянистых, так и среди деревянистых растений (кизильник горизонтальный — Cotoneaster saxatile f. horizontale). Предполагают, что у древесных и кустарниковых растений стелющиеся стебли являются результатом приспособления к неблагоприятным условиям. Много видов со стелющимися стеблями произрастает в субарктических и арктических областях, в горах, а также на океанических побережьях. У травянистых растений стелющиеся или ползучие стебли часто образуются при сильном затенении или избыточном увлажнении (лисохвост коленчатый — Alopecurus geniculatus, полевица побегообразующая — Agrostis stolonizans).
Особым типом стеблей является укороченный стебель. Укороченный стебель имеет сильно сближенные узлы и погружен в почву, поэтому над субстратом образуется розетка листьев (одуванчик лекарственный — Taraxacum officinale, подорожник большой — Plantago major и др.). Цветоносные стебли (цветоносы) у таких растений безлистные, прямостоячие и называются стрелками.
Различаются стебли и по поперечному сечению (рис. 94). Наиболее типичным является цилиндрический стебель. У злаков полый в междоузлиях круглый стебель называется соломиной.
Рис. 94. Виды стеблей по поперечному сечению: 1—цилиндрический; 2—сплюснутый; 3—трехгранный; 4—тупотрехгранный; 5—выгнутотрехгранный; 6—четырехгранный; 7—пятигранный; 8—многогранный; 9—ребристый; 10—бороздчатый; 11—крылатый
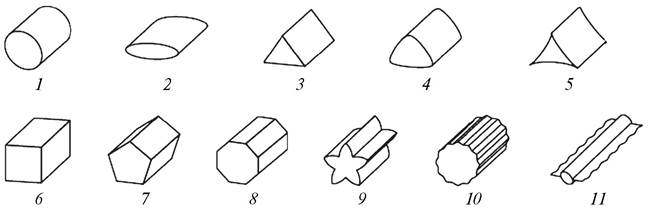
Стебли могут быть трехгранными (виды рода осока — Carex), четырехгранными (сем. Губоцветные — Labiatae), сплюснутыми (мятлик сплюснутый — Poa compressa, опунция — Opuntia), ребристыми (гладыш прусский — Laserpitium prutenicum, хвощ полевой — Equisetum arvense).
Своеобразное строение имеет крылатый стебель (чина лесная — Lathyrus sylvestris, чертополох курчавый — Carduus crispus). У этих растений вдоль круглого стебля на всем протяжении или участками прикреплены листовидные образования. Поперечное сечение такого стебля похоже на рыбий позвонок.
Морфологические особенности стебля имеют важное диагностическое значение и используются при идентификации видов покрытосеменных растений.
Указанные морфологические особенности характерны для стеблей, выполняющих типичные функции. При выполнении дополнительных функций стебель метаморфизируется, а вместе с ним метаморфизируются и листья. Таким образом, следует говорить не просто о метаморфозе стебля, а о метаморфозе побега в целом.
5.3.5. Метаморфозы побега
Метаморфозы побега более разнообразны, чем метаморфозы корня. Под влиянием условий существования может изменяться как побег в целом, так и отдельные его компоненты, в частности лист. Довольно часто метаморфизированные образования надземных органов различного происхождения, выполняющие одни и те же функции, имеют сходное строение. В то же время под влиянием условий существования некоторые образования, происходящие от одного и того же органа, выполняют разные функции и имеют различное анатомическое и морфологическое строение. В связи с понятием метаморфозов надземных органов возникли понятия об аналогичных и гомологичных органах.
Аналогичными называются органы, которые имеют различное происхождение, но выполняют одинаковые функции и имеют сходный внешний вид (морфологическое строение). Например, колючки, усики стеблевого и листового происхождения. Определить в таком случае происхождение метаморфизированного образования можно по расположению их на стебле (по топографии органа). Метаморфозированные образования стеблевого (побегового) происхождения находятся в пазухе листа (боярышник однопестичный — Crataegus monogyna) или располагаются на верхушке побега (груша обыкновенная — Pyrus communis). Метаморфозы листового происхождения располагаются на стеблевом узле (барбарис обыкновенный — Berberis vulgaris).
Гомологичными называются органы, имеющие одинаковое происхождение, но выполняющие разные функции, вследствие чего имеют различное строение. Гомологичны листу колючки листового происхождения (барбарис обыкновенный — Berberis vulgaris), усики листового происхождения (горох посевной — Pisum sativum, чина луговая — Lathyruspratensis),сухие чешуи на корневищах (пырей ползучий — Elytrigia repens, осока мохнатая — Carex hirta), сочные чешуи луковиц (лук репчатый — Allium cepa, нарцисс гибридный — Narcissushybridus) и др.
К метаморфозам побега относятся корневища, луковицы, клубни, столоны, кладодии (или филлокладии), колючки и усики.
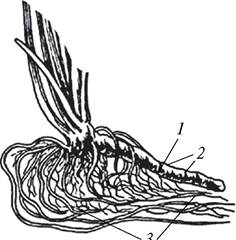
Корневище (rhizoma) — подземный метаморфизированный побег, у которого в отличие от типичного побега листья редуцированы и превращены в сухие (осока мохнатая — Carexhirta) или сочные (сусак зонтичный — Butomus umbellatus) чешуи (рис. 95).
Рис. 95. Плагиотропное сочное корневище сусака зонтичного (Butomus umbeПatus): 1—узел; 2—междоузлие; 3—придаточные корни
По особенностям формирования различают корневища эпигеогенные (копытень европейский — Asarum europaeum, представители родов манжетка — Alchemilla, гравилат — Geum,медуница — Pulmonaria) и гипогеогенные (ландыш майский — Convallaria majalis, грушанка круглолистная — Pyrola rotundifolia и др.) (рис. 96).
Рис. 96. Типы корневищ: А—Г — плагиотропные; Д—ортотропное: А—эпигеогенное корневище гравилата речного (Geum rivale); Б—моноподиальное корневище вороньего глаза обыкновенного (Paris quadrifolia); В—гипогеогенное корневище ландыша майского (Convallaria majalis); Г—симподиальное корневище купены душистой (Polygonatum odoratum); Д—ортотропное корневище веха ядовитого (Cicuta virosa): 1—стеблевой узел; 2—междоузлие; 3—боковые почки
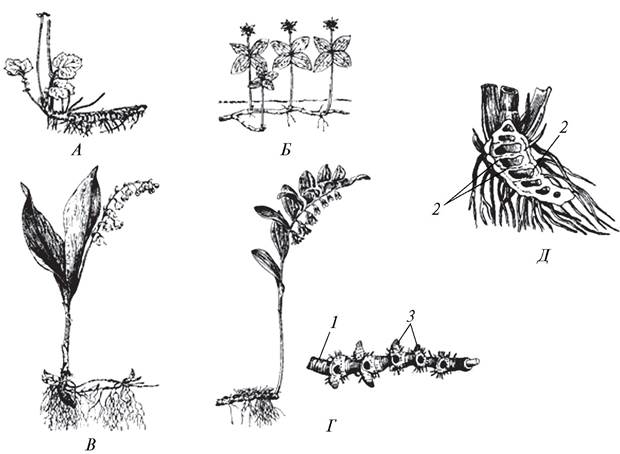
Эпигеогенное (от греч. epi — над, geo — земля, genos — род, происхождение) корневище развивается из почки, расположенной на надземном побеге, и занимает подземное положение в результате контрактильности корней, которые «втягивают» корневище в почву, или засыпания его листовым опадом.
Гипогеогенное (от греч. hypo — под, внизу, geo — земля, genos — род, происхождение) корневище развивается из почки, расположенной на подземном побеге. На стебле гипогеогенного корневища листья редуцированы и представлены пленчатыми чешуями.
По направлению роста корневища бывают плагиотропные — горизонтально распростертые (гравилат речной — Geum rivale) и ортотропные (вех ядовитый — Cicuta virosa), растущие вертикально по направлению силы тяжести Земли.
Корневища могут ветвиться моноподиально (вороний глаз четырехлистный — Paris quadrifolia) и симподиально (купена лекарственная — Polygonatum odoratum). По консистенциикорневища бывают сухими (пырей ползучий Elytria repens) и сочными (аир обыкновенный — Acorus calamus, ирис безлистный — Iris aphylla).
Луковица (bulbus) — метаморфизированный побег с укороченным, уплощенным стеблем (донцем) и чешуевидными листьями, имеющими разное происхождение и строение (рис. 97). В зависимости от места образования на растении луковицы бывают подземными и надземными. Надземные луковицы имеют небольшие размеры, их часто называют бульбочками, или луковичками. Луковички могут формироваться в пазухах листа (лилия тигровая — Lilium tigrinum, зубянка луковичная — Dentaria bulbifera) или в соцветии (чеснок — Allium sativum,мятлик луковичный — Poa bulbosa, лук огородный — Allium oleraceum).
Рис. 97. Типы луковиц: А—Б — туникатная(покровная) луковица лука репчатого (Allium cepa): А—схема строения луковицы; Б—внешний вид луковицы; В—черепитчатая луковица лилии кудреватой (Lilium martagon); Г — сложная луковица чеснока (Allium sativum): 1— донце (метаморфизированный укороченный стебель); 2—придаточные корни; 3—сухие чешуи; 4—сочные чешуи; 5—зачаточная почка; 6—основания листьев; 7—дочерние луковички; 8—общая кроющая сухая чешуя
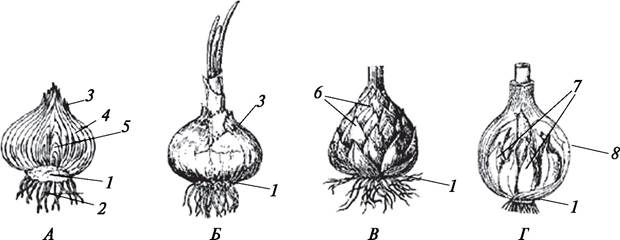
По особенностям расположения чешуй различают луковицы туникатные (пленчатые, концентрические) и черепитчатые. Туникатные луковицы образованы сросшимися концентрически расположенными на донце — уплощенном стебле — чешуями. Число чешуй в луковице варьирует от одной (чеснок — Allium sativum) до многих (лук репчатый — Allium cepa). У черепитчатых луковиц чешуи расположены в шахматном порядке. По степени сложности луковицы бывают простыми и сложными. В сложной луковице (чеснок) под общей сухой чешуей располагается много простых луковичек.
Поскольку луковицы являются метаморфизированными побегами, их можно различать и по типам ветвления. У симподиальных луковиц (тюльпан гибридный — Tulipa hybrida, рябчик королевский — Fritillaria imperialis, лук репчатый — Allium cepa) цветонос формируется из верхушечной почки, а возобновление (образование дочерних луковиц) происходит из пазушных почек. У моноподиальных луковиц (подснежник снежный — Galanthus nivalis, нарцисс гибридный — Narcissus hybridus) возобновление идет из верхушечной почки, а цветоносы — из пазушных почек.
По продолжительности жизни луковицы бывают многолетними (гипе- аструм гибридный — Hyppeastrum hybridum, нарцисс) и однолетними (тюльпан, лук). Однолетние луковицы ежегодно отмирают, а вместо отмершей (материнской) луковицы образуется замещающая луковица и луковицы-детки. Растения, у которых луковицы ежегодно отмирают, следует считать не многолетниками, а вегетативными малолетниками.
Вегетативными малолетниками являются и клубнелуковичные растения (шпажник черепитчатый — Gladiolus imbricatus, шафран посевной — Crocus sativus).
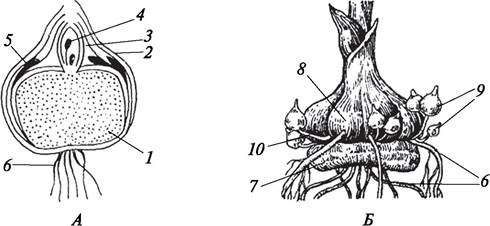
Клубнелуковица (bulbotuber) в отличие от луковицы формируется преимущественно за счет разрастания и уплощения стебля (рис. 98). Листья на клубнелуковицах чешуйчатые, они надежно прикрывают верхушечную и пазушные почки. Клубнелуковицы, как и луковицы, могут быть как однолетними (шафран весенний — Crocus vernus, гладиолус гибридный — Gladiolus hybridus), так и многолетними (безвременник осенний — Colchicum autumnale).
Рис. 98. Клубнелуковицы гладиолуса (Gladiolus hybridus): А—схема строения клубнелуковицы; Б—внешний вид формирующихся клубнелуковиц и клубнелуковичек: 1— метаморфизированный стебель; 2—кроющая чешуя; 3—зачаточные листья; 4—зачаточная цветочная почка; 5—редуцированные чешуевидные листья; 6—придаточные корни; 7—отмирающая старая клубнелуковица; 8—формирующаяся замещающая клубнелуковица; 9—клубнелуковички-детки; 10—столон
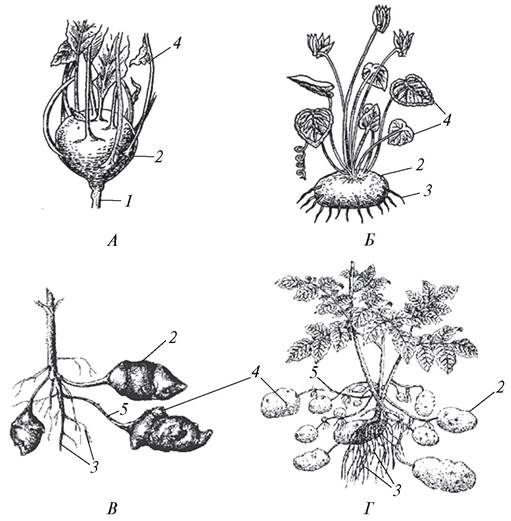
Клубни (tuber) определяют, как метаморфизированные подземные побеги, сильно утолщенные за счет образования большого количества паренхимной ткани. Однако у некоторых растений клубни или мелкие образования — клубеньки возникают в надземной части растения. Мощный надземный клубень образуется в результате разрастания стебля у разновидности капусты огородной — кольраби (Brassica oleracea var. longeloides).
У ряда тропических эпифитных орхидей основание стебля сильно разрастается и превращается в туберидий (кеттлея — Cattleya, максиллярия — Maxillaria). У горца живородящего (Polygonum vivipara) клубеньки образуются в соцветиях, а у чистяка весеннего (Ficaria verna) — в пазухах листьев.
Подземные клубни могут иметь различное происхождение. Если у картофеля (Solanum tuberosum), топинамбура (Helianthus tuberosus) это метаморфизированные побеги, о чем свидетельствует наличие почек (верхушечных и пазушных), стеблевых узлов (бровка), междоузлий, редуцированных до едва заметных чешуек листьев, то у цикламена индийского (Cyclamen indica) подземный клубень образован только частью стебля — гипокотилем (рис. 99).
Рис. 99. Клубни стеблевого происхождения: А—надземный клубень капусты кольраби (Brassica oleracea var. gongylodes); Б—подземный клубень цикламена индийского (Cyclamenindica); В—подземные клубни топинамбура (Helianthus tuberosus); Г—подземные клубни картофеля (Solanum tuberosum): 1—типичный стебель; 2—стебель клубневидно утолщенный; 3—придаточные корни; 4—почки; 5—столоны
Столоны — безлистные образования, фактически единственное сильно удлиненное междоузлие подземного побега, несущее на конце клубень (картофель) или луковицу (лук угловатый — Allium angulosum). Столоны обычно плагиотропы, однако у дикорастущих тюльпанов и у культурных, если их ежегодно не выкапывают, образуются ортотропные столоны, и дочерние луковицы постепенно сильно заглубляются. Это приводит к измельчению луковиц и к быстрому вырождению сорта.
Метаморфозы надземных побегов, как и подземных, многообразны (рис. 100). У растений засушливых местообитаний побеги могут превращаться в кладодии (от греч. cladus — ветвь) — листоподобно уплощенные образования (шлюмбергера, или «декабрист», — Schlumbergera sp.), или филлокладии (иглица — Ruscus, спаржа — Asparagus). Термины кладодии и филлокладии некоторые ботаники рассматривают как синонимы, другие придают им самостоятельное значение и указывают, что для кладодий характерен длительный рост, а рост филлокладий ограничен. И кладодии, и филлокладии повернуты ребром к солнцу, что имеет важное приспособительное значение (уменьшение испарения, защита от перегревания). Кладодии и филлокладии всегда располагаются в пазухах чешуйчатых «листьев», что подтверждает их побеговое (стеблевое) происхождение. Об этом же свидетельствует образование цветков. У иглиц на многолетних кладодиях находятся довольно длинные цветоносы, на концах которых ежегодно формируются раздельнополые цветки.
Рис. 100. Метаморфозы надземных побегов: А—иглица колючая (Ruscus aculeatus); Б—огурец посевной (Cucumis sativus); В—переступень белый (Bryonia alba); Г— боярышник(Crataegus sp.): 1—типичный стебель; 2—филлокладий — листоподобно уплощенный боковой побег; 3—редуцированный пленчатый лист; 4—цветок; 5— усики стеблевогопроисхождения; 6—колючки стеблевого происхождения
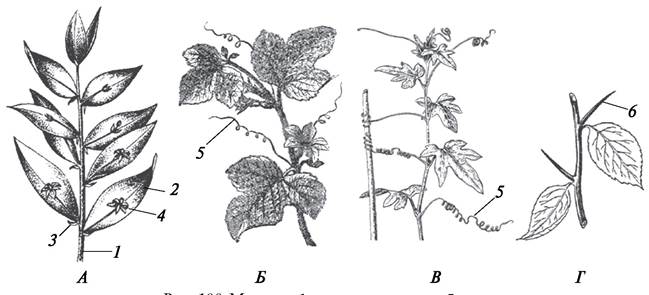
Колючки стеблевого происхождения характерны для ряда деревьев (груша обыкновенная — Pyrus communis) и кустарников (боярышник однопестичный — Crataegus monogina). У формирующихся молодых побегов, из которых впоследствии разовьются колючки, на стебле видны зачаточные, недоразвитые листья. Они расположены так же, как и типичные листья. С возрастом колючка одревесневает, а зачаточные листья абортируются, т. е. полностью исчезают. Колючки стеблевого происхождения или выходят из пазухи листа, или развиваются из верхушечной почки. Они выполняют защитную функцию.
Усики стеблевого происхождения, как и колючки, или выходят из пазухи листа, или заканчивают каждый метамер симподиально ветвящегося стебля (огурец посевной — Cucumis sativus, виноград культурный — Vitis vinifera). Усики могут быть простыми или разветвленными, основная их функция опорная.
Таким образом, метаморфозы побега разнообразны. Метаморфизирован- ные побеги выполняют различные функции, в том числе функции сохранения и размножения вида (клубни, корневища, луковицы, клубнелуковицы).