МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 3. ТКАНИ РАСТЕНИЙ
3.4. Анатомо-морфологические и функциональные особенности основных (паренхимных) тканей
Паренхима — это постоянная ткань, составляющая большую часть тела растения. Она занимает пространство между всеми другими постоянными, более специализированными тканями, поэтому ее можно обнаружить во всех органах растения. В связи с этим паренхиму иногда называют основной тканью. Находясь в тургесцентном состоянии, клетки паренхимы оказываются плотно упакованными, что обеспечивает дополнительную опору органам растений, особенно травянистых. При потере воды клетки спадаются и растения вянут. Паренхима — это единственная из постоянных тканей, которая может дедифференцироваться и восстанавливать меристематическую активность. Благодаря этому происходит заживление ран при повреждении растений, а также регенерация органов при вегетативном размножении. Паренхимная ткань чаще всего первична по происхождению. Она образуется из основной меристемы апексов корня и стебля. Ее клетки тонкостенные, обычно изодиаметрической, иногда вытянутой формы, стенки имеют простые поры и первичное строение. Иногда клеточные стенки паренхимы утолщаются и одревесневают. Такая паренхима называется склеренхиматозной. Паренхима хорошо развита в первичной коре стебля и корня, в сердцевине, сердцевинных лучах, мезофилле листьев, в сочных плодах.
Паренхима является полифункциональной тканью. Клетки паренхимы имеют различное строение и в зависимости от положения в теле растения выполняют разные функции. Основные (паренхимные) ткани являются преимущественно питающими тканями, хотя по своему существу могут выполнять многие функции, начиная c ассимиляционной, заканчивая выделительной. Независимо от положения в теле растения паренхима обеспечивает транспорт веществ в горизонтальном направлении по клеточным стенкам (апопласту) и цитоплазматическим путям (симпласту).
3.4.1. Типы и функции паренхимных тканей
В зависимости от выполняемой функции и строения клеток различают ассимиляционную (хлорофиллоносная ткань, или хлоренхима), воздухоносную (аэренхима), запасающую, трансфузионную, древесинную (склеренхиматозную) и лубяную паренхиму.
Ассимиляционная паренхима (хлоренхима) осуществляет фотосинтез (рис. 26). Именно в этой ткани, несмотря на ее относительно простое строение, происходит фиксация и преобразование солнечной энергии, а также синтез основной массы органических веществ на Земле. Она состоит из тонкостенных живых клеток, содержащих хлоропласты, поэтому нередко ее называют хлоренхимой. Хлоренхима присутствует во всех зеленых частях растения.
Рис. 26. Схема строения клеток разных типов хлоренхимы: А—столбчатая (палисадная); Б—губчатая (рыхлая); В—складчатая: 1—хлоропласты; 2—межклетники; 3— впячивание клеточной стенки
Чаще всего она залегает прямо под эпидермой. Близкое к поверхности расположение ассимиляционной ткани придает зеленую окраску органам растения. Подавляющая часть ассимиляционной ткани находится в листьях, там она составляет мезофилл (от греч. mesos — средний и phyllon — лист). Форма клеток хлоренхимы различна, соответственно им выделяют три ее типа: столбчатую, или палисадную; губчатую, или рыхлую; складчатую.
Клетки столбчатой хлоренхимы имеют таблитчатую форму, они плотно прилегают друг к другу, содержат большое число активных хлоропластов. В этой хлоренхиме интенсивно идет процесс фотосинтеза, активно осуществляется циклоз — круговое или струйчатое движение цитоплазмы. Губчатая хлоренхима состоит из более или менее округлых клеток с довольно большими межклетниками. Она расположена глубже, чем столбчатая хлоренхима, процессы фотосинтеза в ней протекают менее активно, зато хорошо развитая система межклетников обеспечивает интенсивное перемещение газов и паров воды. Дифференциацию на столбчатую и губчатую хлоренхиму можно наблюдать только у листьев, имеющих дорзовентральное строение и находящихся в условиях яркого освещения. В листьях сосны, которые обычно живут несколько лет и попадают в условия сезонных колебаний температуры и влажности, развивается складчатая хлоренхима. Она сформировалась в процессе эволюции как адаптивный признак, поскольку игольчатая форма листа резко снижает испаряющую поверхность, но должна обеспечить нормальный фотосинтез. В связи с этим в паренхиме мезофилла листьев сосны оболочки клеток врастают внутрь, образуя складки. В результате значительно увеличивается протяженность прилегающего к оболочке слоя цитоплазмы с хлоропластами и общая фотосинтезирующая поверхность.
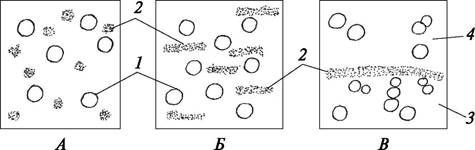
Запасающая паренхима может развиваться в разных органах и частях растения. В ней откладываются избыточные в данный период развития продукты метаболизма: белки, углеводы (сахара, инулин, крахмал), липиды (рис. 27). Они могут накапливаться в жидком (сахара, масла) или твердом (крахмал, белки и др.) состоянии. Обычно запасные вещества накапливаются внутри клетки, но могут откладываться и в клеточной стенке. Запасающие ткани хорошо развиты в плодах, метаморфизированных подземных органах — корне - плодах, корневых шишках и др., а также в сердцевине и коре стеблей. Местом хранения резервных веществ может быть также паренхима проводящих тканей. В семенах запасные питательные вещества сосредоточены в эндосперме (зонтичные, злаки), семядолях зародыша (бобовые) или в окружающем зародыш перисперме (гвоздичные, маревые). У финиковой пальмы гемицеллюлоза, являющаяся запасным продуктом, откладывается в стенках клеток эндосперма.
Рис. 27. Клетки запасающей паренхимы с различными веществами: А—кристаллы инулина в клетках корневых клубней георгины садовой (Dahlia variabilis); Б— крахмальные зерна в клетках черешка пеларгонии зональной (Pelargonium zonale); В—алейроновые зерна в семенах клещевины обыкновенной (Ricinus communis): 1— кристаллы инулина; 2—сложные крахмальные зерна; 3—алейроновое зерно; 4—оболочка алейронового зерна; 5—кристаллоид белка; 6—глобоид белка
У растений засушливых мест обитания (ксерофитов) развивается паренхима, запасающая воду (кактусы, растения сем. Толстянковые). Развитой водоносной тканью снабжены клубневидные вздутия стеблей многих эпифитных орхидных, а также метаморфизированные корни некоторых лилейных (хлорофитум, спаржа). В тонкостенных клетках водозапасающей паренхимы находится много слизи, которая и удерживает влагу. При подсыхании растения водоносные клетки передают воду главным образом ассимиляционным тканям.
Подавляющее большинство запасных питательных веществ, которые являются резервными, на определенных этапах развития растения вновь вовле - каются в процессы метаболизма.
Воздухоносной паренхимой, или аэренхимой, называют паренхиму с сильно развитыми межклетниками. Основная функция этой ткани — вентиляция вегетативных органов, которые находятся в условиях пониженного содержания кислорода. Она характерна для водно - болотных растений, но встречается и у сухопутных видов (например, у ситников, произрастающих в условиях пониженной аэрации почвы). В межклетниках этой ткани содержится значитель ное количество газа, который отличается по своему составу от воздуха, что позволяет дополнительно снабжать лист кислородом или углекислым газом. Кроме того, аэренхима водных растений обеспечивает плавучесть растения. Аэренхима развивается в корне, стебле, листьях, иногда образуется в плодах (снежноягодник) (рис. 28).
Рис. 28. Аэренхима: А—в черешке листа кувшинки белой (Nymphaea alba); Б—в стебле ситника развесистого (Juncus effusus); В—в стебле рдеста блестящего (Potamogeton lucens); Г—в корне жюссеи ползучей (Jussiaea repens): 1—кутикула; 2—эпидермис; 3—клетки воздухоносной паренхимы (аэренхимы); 4—воздухоносные полости; 5—центральный цилиндр
Трансфузионная (передаточная) паренхима формируется в листьях хвойных растений между сосудисто волокнистыми пучками и эндодермой. Она со стоит из живых клеток с первичными оболочками и участвует в проведении веществ. Функционально клетки трансфузионной паренхимы сходны с альбуминовыми клетками (клетками Страсбургера), которые характерны для флоэмы голосеменных растений. Этот вид паренхимы вместе с трансфузионными трахеидами — мертвыми водопроводящими клетками — входит в состав трансфузионной ткани. Она способствует передаче как продуктов фотосин теза от хлоренхимы листа к проводящим пучкам, так и воды от сосудисто волокнистых пучков к хлорофиллоносной паренхиме.
Особыми типами паренхимных тканей являются древесинная и лубяная паренхима.
Древесинная (склеренхиматозная) паренхима — это живая ткань с утолщенными клеточными стенками клеток, которая формирует во вторичной ксилеме вертикальные ряды (вертикальную, или тяжевую паренхиму). Оболочки древесинной паренхимы слабо одревесневшие с многочисленными простыми порами. Тяжевая паренхима выполняет функцию запаса питательных веществ. Основным запасным веществом является крахмал. Особенно хорошо тяжевая паренхима выражена у древесных растений.
В зависимости от расположения по отношению к сосудам ксилемы у них различают апотрахеальную и паратрахеальную паренхиму.
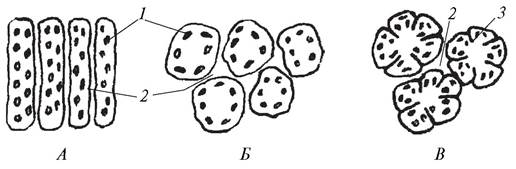
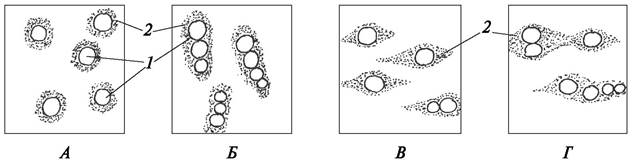
Апотрахеальная паренхима не связана с сосудами. Она может быть диффузной, терминальной или метатрахеальной (рис. 29). Клетки диффузной паренхимы беспорядочно разбросаны среди элементов годичного кольца (кольца прироста). Апотрахеальная паренхима, которая образуется в конце годичного прироста (осенью), называется терминальной. Апотрахеальная паренхима в виде коротких полосок, расположенных тангентально к поверхности, называется метатрахеальной.
Рис. 29. Апотрахеальная паренхима. Схема расположения клеток тяжевой древесинной паренхимы в древесине (вторичной ксилеме) лиственных деревьев: А— диффузная; Б—метатрахеальная; В—терминальная: 1—сосуды; 2—тяжевая (вертикальная) паренхима; 3—наружный слой поздней древесины (граница годичного прироста); 4—древесина нового годичного кольца (весенняя древесина)
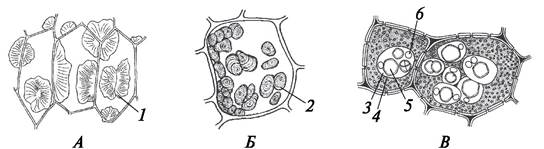
Паратрахеальная паренхима располагается вокруг сосуда и образует полную или частичную обкладку. Она бывает вазицентрической, крыловидной, сомкнуто-крыловидной (рис. 30). Вазицентрическая паренхима формирует полную обкладку вокруг сосуда. Крыловидная паренхима примыкает лишь к отдельным сосудам, образуя своеобразный крыловидный выступ. Если крыловидные обкладки сосудов соединяются между собой, паратрахеальная паренхима называется сомкнуто-крыловидной.
Рис. 30. Паратрахеальная паренхима. Схема расположения клеток тяжевой древесинной паренхимы в древесине (вторичной ксилеме) лиственных деревьев: А, Б— вазицентрическая паренхима; В—крыловидная паренхима; Г—сомкнутокрыловидная паренхима: 1—сосуды; 2—тяжевая (вертикальная) паренхима
Лубяная паренхима входит в состав вторичной флоэмы. Так же, как и древесинная паренхима, она является тяжевой (вертикальной), непосредственно примыкает к ситовидным элементам и взаимодействует с ними. Полагают, что живые клетки лубяной паренхимы вырабатывают и передают в ситовидные трубки ферменты, необходимые для ускорения передвижения продуктов фотосинтеза. Кроме того, в лубяной паренхиме могут откладываться запасные питательные вещества, а также продукты (экскреты), выводимые из процесса метаболизма. Новообразование клеток лубяной паренхимы приводит к дилатации (разрастанию) луба, а утолщение клеточной стенки паренхимных клеток завершается образованием склереид, что способствует увеличению механической прочности стебля и корня. Клетки лубяной паренхимы могут дедифференцироваться и образовывать клетки феллогена, из которого впоследствии формируются внутренние перидермы.
3.5. Анатомо-морфологические и функциональные особенности механических (арматурных) тканей
Механические ткани в теле растения выполняют опорную функцию, придавая определенную конфигурацию отдельным органам. Чаще всего они играют роль арматуры, располагаясь среди других тканей и придавая им в сочетании с остальными тканями растения дополнительную прочность. Механические ткани обеспечивают сопротивление растения статическим (сила тяжести) и динамическим (порывы ветра, удары дождя и т. д.) нагрузкам. Общим и главным их признаком является утолщение клеточных оболочек и тесная сомкнутость конструктивных элементов. В процессе эволюции растений механические ткани возникли в связи с совершенствованием ветвления и развитием вегетативной массы растения.
В зависимости от времени образования в онтогенезе растений различают первичные и вторичные механические ткани: первичные образуются из основной меристемы, вторичные — из камбия.
Различают два типа механических тканей — колленхиму и склеренхиму.
3.5.1. Колленхима как тип механической ткани
Колленхима (от греч. colla — клей и enchyme — налитое) представляет собой ткань из живых клеток длиной 1—2 мм с неравномерно утолщенными первичными клеточными стенками. Утолщения возникают на первичной оболочке преимущественно за счет отложения небольшого количества молекул целлюлозы и обильного отложения пектиновых веществ. Они никогда не пропитываются лигнином и не одревесневают, сохраняют упругость, оставаясь живыми, как правило, в течение всей жизни растения. В связи с тем, что оболочки клеток имеют первичное строение, они способны к растяжению и не препятствуют росту других клеток, находящихся рядом. Именно поэтому колленхима является опорной тканью молодых растущих в длину органов. Клетки колленхимы вытягиваются параллельно длинной оси органа, в котором закладывается эта ткань. Важной особенностью колленхимы является то, что она выполняет функцию опоры только в состоянии тургора. В противном случае при потере растением воды тонкие участки клеточных оболочек спадаются, побеги теряют упругость, растения увядают.
Колленхимные ткани имеют первичное происхождение, часто образуются из внешних слоев апикальной меристемы побега (в корне колленхима не образуется), и поэтому расположены (у двудольных) под эпидермисом в первичной коре. При этом колленхима образуется в виде кольца или сосредоточена в ребрах (в стеблях яснотки, черешках листа подорожника, сельдерея), что, несомненно, повышает прочность органа. Присутствие колленхимы в черешке листа у так называемых компасных растений или под соцветием (у подсолнечника) позволяет этим растениям менять ориентацию листьев или соцветий в пространстве вслед за положением солнца. В листьях некоторых двудольных растений колленхима может располагаться над и под жилкой и служит опорой проводящим пучкам. У однодольных колленхима находится в узлах злаков и предохраняет растения от полегания.
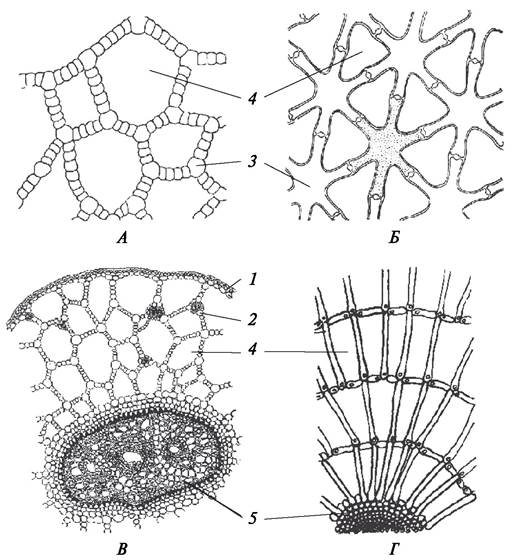
В зависимости от характера утолщения клеточных стенок различают три типа колленхимы — уголковую, пластинчатую и рыхлую (рис. 31).
Рис. 31. Типы колленхимы: А—пластинчатая колленхима в стебле однолетнего побега дуба черешчатого (Quercus robur); Б—уголковая колленхима в черешке листа бегонии королевской (Begonia rex); В—рыхлая колленхима в стебле красавки белладонны (Atropa belladonna): 1—эпидермис; 2—утолщение клеточной стенки; 3— полость клетки; 4—межклетники; 5—формирующаяся перидерма
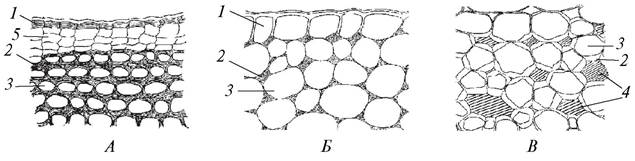
В уголковой колленхиме утолщения сосредоточиваются по углам клеток и контактируют с утолщениями в соседних клетках. В результате клетки выглядят многогранными (5—6-гранными), а совокупность утолщенных оболочек соседних клеток имеет вид 3—5-угольников. Утолщенные участки целлюлозных оболочек выглядят блестящими. Уголковая колленхима хорошо развита в черешках бегонии, ревеня, свеклы, тыквы, щавеля, гречихи и других растений.
В пластинчатой колленхиме сильно утолщены тангентальные (параллельные поверхности органа) клеточные стенки. В таких клетках на ранних этапах развития ткани радиальные стенки остаются более тонкими. Пластинчатая колленхима представляет более прочную ткань, чем уголковая. Она развивается в стеблях подсолнечника, бузины, баклажана, астранции.
Рыхлая колленхима начинает формироваться как уголковая, однако в результате расхождения клеток образуются большие межклетники. Они играют роль схизогенных вместилищ, в которых накапливаются слизистые выделения, или выполняют воздухоносную функцию. Полость клетки при этом сильно уменьшается и почти не видна. Такой тип колленхимы можно наблюдать в черешках лопуха, подбела, в цветоносах растений из семейства Лилейные (валоты, лилии, чеснока).
Клетки колленхимы, располагаясь под эпидермисом в периферической части первичной коры, могут содержать хлоропласты и выполнять не только опорную функцию, но и функцию фотосинтеза. Со временем оболочки клеток колленхимы изредка утолщаются, одревесневают и колленхима превращается в склеренхиму. Иногда клетки колленхимы могут терять утолщения и превращаться в запасающую паренхиму.
3.5.2. Склеренхима как тип механической ткани
Склеренхима (от греч. scleros — твердый и еnchyme — налитое) — более прочная механическая ткань. Она встречается во всех органах растения и располагается не только под эпидермисом, но и в других частях растения. В отличие от колленхимы клетки склеренхимы имеют равномерно утолщенные клеточные стенки, в которых располагаются немногочисленные простые поры (рис. 32). Как правило, у клеток склеренхимы образуются вторичные клеточные оболочки. Оболочки зрелых клеток склеренхимы пропитываются лигнином и одревесневают, а живое содержимое клеток постепенно отмирает. Таким образом, зрелые клетки склеренхимы в отличие от колленхимы чаще всего мертвые. Такие клетки не способны растягиваться, поэтому окончательное их созревание происходит после того, как закончится рост окружающих клеток. Оболочки склеренхимных клеток обладают высокой прочностью, близкой к прочности стали. Они хорошо выдерживают динамические нагрузки и не испытывают остаточной деформации, поэтому обеспечивают прочность на сжатие, разрыв, изгиб. Высокая прочность на разрыв означает возможность значительного растяжения без повреждения органов растения, а прочность на сжатие обеспечивает достаточное сопротивление изгибу.
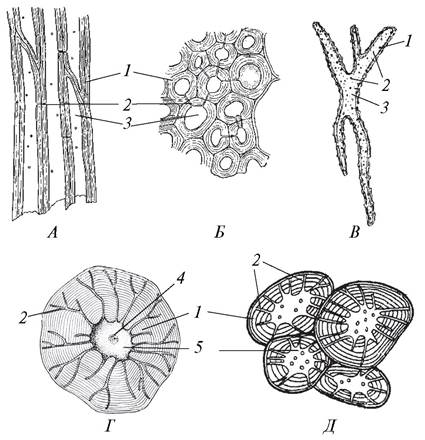
Рис. 32. Склеренхима: А, Б—продольный и поперечный разрез волокон в стебле пеларгонии зональной (Pelargonium zonale); В—астросклереида в листовой пластинке кувшинки белой (Nymphaea alba); Г—склереида из скорлупы кокосового ореха (Cocos nucífera); Д—каменистые клетки из мякоти плода груши (Pyrus communis): 1— вторичная клеточная оболочка; 2—поровые каналы; 3—полость клетки; 4—ядро; 5—центральная часть клетки
Иногда клетки склеренхимы не одревесневают, что характерно для прядильных растений (волокна льна, кенафа, кендыря). У текстильных культур волокна остаются целлюлозными, сохраняют гибкость и эластичность. В редких случаях, благодаря многочисленным разветвленным простым порам, клетки склеренхимы сохраняют живое содержимое (каменистые клетки в плодах).
По происхождению различают склеренхиму первичную и вторичную. Первичная склеренхима чаще всего возникает из меристем апекса побега и располагается в стебле под эпидермисом (у однодольных растений). Если она образуется из прокамбиальных тяжей, то находится в обкладке проводящих пучков. Первичная склеренхима может формироваться из перицикла. Перициклическая склеренхима характерна для стеблей кирказона, купены, льна.
Вторичная склеренхима образуется из камбия и входит в состав обкладки проводящих пучков или диффузно разбросана среди паренхимных тканей различных частей растения.
Склеренхима подразделяется на волокна и склереиды (см. рис. 32).
Волокна представлены сильно вытянутыми прозенхимными клетками с заостренными концами (скошенными поперечными стенками). Клетки растут скользящим (интрузивным) ростом, перекрывают друг друга, что повышает механическую прочность ткани. Волокна имеют толстые клеточные стенки и очень узкую полость клетки. Их длина в среднем 1—4 мм, но у текстильных культур значительно больше: до 60 мм у льна, до 350 мм у рами. Чем длиннее волокна, тем выше их технические показатели. Номер волокна у льна отражает его длину: чем выше номер, тем длиннее волокно. Волокна, расположенные во флоэме (или в лубе, как часто именуют флоэму), называются лубяными. Склеренхимные волокна, которые находятся в ксилеме (древесине) называются древесинными, или либриформом. Они короче лубяных (не более 2 мм) и всегда одревесневают. Предполагают, что в процессе эволюции волокна либриформа образовались из лестничных трахеид — водопроводящих элементов ксилемы.
Волокна могут также входить в состав других тканей, располагаясь целыми группами или поодиночке. У многих растений, особенно у однодольных, волокна составляют механическую обкладку проводящих пучков. Элементы механических тканей, входящие в состав сосудисто-волокнистых пучков, характеризуются относительно короткой длиной и более тонкой оболочкой по сравнению с механическими элементами, которые находятся вблизи поверхности органа.
Склереиды — это клетки, близкие по форме к паренхимным, с толстыми вторичными слоистыми стенками и простыми разветвленными порами. Содержимое их обычно отмирает, но иногда сохраняется, как в плодах груши и айвы. У этих растений при созревании плодов клетки раздревесневают и становятся тонкостенными паренхимными запасающими клетками. Обычно наиболее богаты склереидами кора (клюква), сердцевина, флоэма осевых органов (хинное дерево), а также плоды и семена (бобовые), листья. Склереиды в разных органах растений имеют различную форму. Склереиды округло-эллиптической формы называются брахисклереидами (от греч. brachys — короткий), или каменистыми клетками. Они обычно располагаются группами и в большом количестве имеются в незрелых плодах груши, айвы. Астросклереиды (от греч. astron — звезда) характеризуются наличием вытянутых отростков клеток. Они чаще встречаются в толстых кожистых листьях (фикус, камелия, инжир), в черешках некоторых растений (кувшинка, кубышка) и обеспечивают их устойчивость к разрывам. Склереиды, похожие на берцовую кость, называют остеосклереидами (от греч. osteon — кость).
Склереиды могут встречаться также поодиночке, в таком случае их называют идиобластами. Идиобласты формируются в листьях инжира, камелии. Во внутриплоднике (эндокарпии) сочных плодов вишни, сливы, грецкого ореха, во внеплоднике (экзокарпии) сухих плодов фундука, лещины идиобласты образуют прочную механическую структуру — косточку.
Ф. Габерландт предложил называть всю совокупность толстостенных одревесневших клеток растений стереомом (от греч. stereos — твердый, крепкий). Степень развития механических тканей во многом зависит от местообитания растений. Она невелика у гигрофитов — растений водной среды, и значительна у ксерофитов — растений засушливых местообитаний.
В теле растения механические ткани имеют определенную локализацию: в корнях они располагаются в центральной части, а в стеблях — в виде кольца на периферии. Действие на побег различных быстро меняющихся и разнонаправленных нагрузок (порывы ветра, капли дождя, вытаптывание животными) требует более эффективного расположения опорных тканей. Такое расположение в надземных органах, по представлениям С. Швенденера, соответствует принципу двутавровой балки в строительных конструкциях и обеспечивает наибольшую прочность органа на изгиб, растяжение и излом. Что касается корня как подземного органа, то его положение в пространстве поддерживается еще и почвой, поэтому механические ткани корня обеспечивают прежде всего прочность на сжатие. Необходимо отметить, что строение органов растения соответствует принципу достижения прочности при экономичной затрате материала. Известный русский ботаник В. Ф. Раздорский, изучавший строительно-механические принципы в конструкции растений, сравнивал их с железобетонными сооружениями, в которых арматуру составляют механические ткани, а роль заполнителя — все прочие живые ткани. Конструктивные особенности растений учитываются в одном из разделов бионики — науки, решающей задачу создания совершенных технических систем.