Молекулярная биология клетки - Том 1 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
Введение в биологию клетки
Малые молекулы, энергия и биосинтез
Питательные вещества и источники энергии клетки
2.3.1. Молекулы питательных веществ, расщепляясь в три этапа, образуют АТР
Белки, липиды и полисахариды, составляющие большую часть нашей пищи, должны расщепиться на меньшие по размеру молекулы, для того чтобы клетки могли их использовать. Ферментативный распад, или катаболизм, этих молекул можно подразделить на три стадии (рис. 2-18). Сначала мы в общих чертах охарактеризуем все эти стадии, а затем обсудим две из них более подробно.
На стадии 1 крупные молекулы полимеров распадаются на мономерные субъединицы: белки на аминокислоты, полисахариды на сахара, а жиры на жирные кислоты и холестерол. Этот предварительный процесс, называемый пищеварением, осуществляется главным образом вне клеток под действием ферментов, секретируемых в полость пищеварительного тракта. На стадии 2 образовавшиеся небольшие молекулы поступают в клетки и подвергаются дальнейшему расщеплению в цитоплазме. Большая часть углеродных и водородных атомов Сахаров превращается в пируват, который, проникнув в митохондрии, образует там ацетильную группу химически активного соединения ацетилкофермента А (ацетил-СоА) (рис. 2-19). Большое количество ацетил-СоА образуется также при окислении жирных кислот. На стадии 3 происходит полное расщепление ацетильной группы ацетил-СоА до СО2 и Н2О. Именно на этой заключительной стадии образуется большая часть АТР.
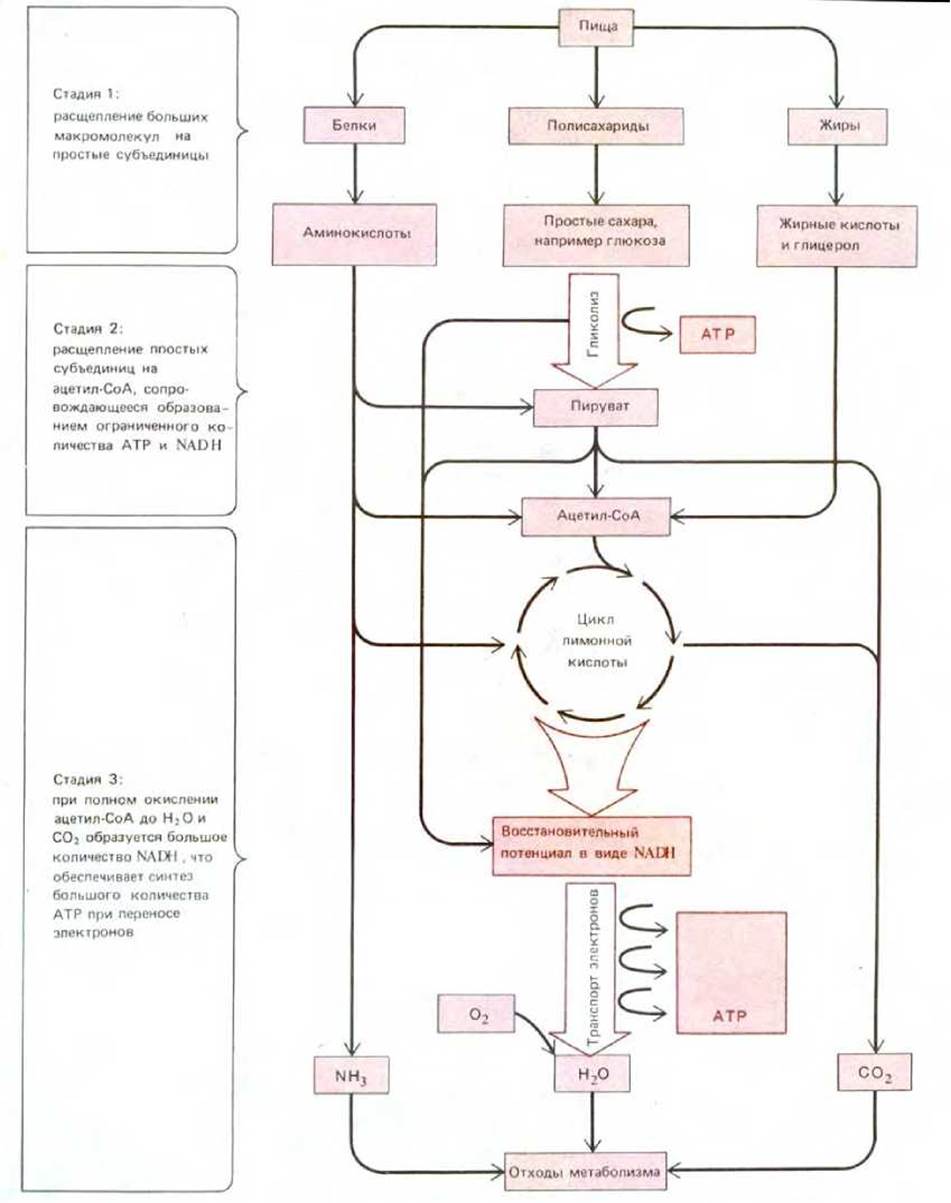
Рис. 2-18. Упрощенная схема трех стадий катаболизма, ведущих от молекул пищевых Веществ к продуктам распада. В этой серии реакций образуется АТР, использующийся затем в биосинтетических реакциях и других энергозависимых процессах.
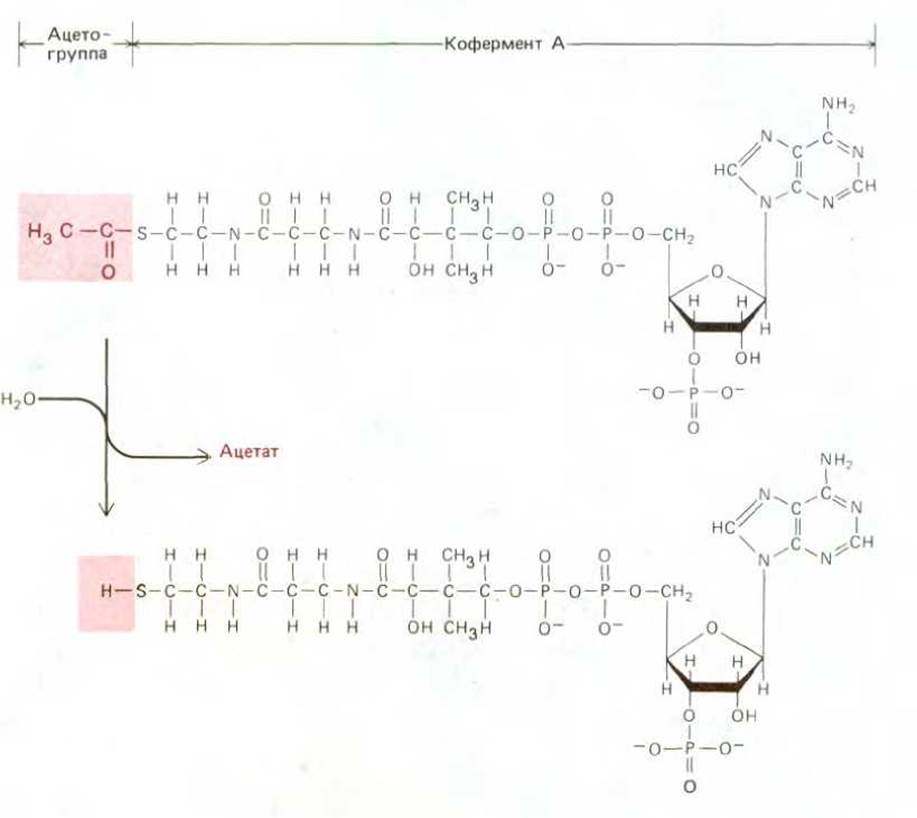
Рис. 2-19. Строение ключевого промежуточного продукта метаболизма - кофермента А (ацетил-СоА). Ацетильные группы, образующиеся на стадии 2 катаболизма (см. рис. 2-18), ковалентно связаны с коферментом А (СоА).
В серии сопряженных химических реакций больше половины той энергии, которую, согласно теоретическим расчетам, можно извлечь из углеводов и жиров при окислении их до Н2О и СО2, используется для осуществления энергетически невыгодной реакции Рi + ADP -> АТР. Поскольку остальная часть энергии, высвобождающейся при окислении, выделяется клеткой в виде тепла, результатом образования АТР является общее возрастание неупорядоченности Вселенной, что полностью соответствует второму закону термодинамики.
Благодаря образованию АТР энергия, первоначально извлеченная путем окисления из углеводов и жиров, преобразуется в более удобную концентрированную форму химической энергии. В растворе, находящемся во внутриклеточном пространстве типичной клетки, имеется примерно 1 млрд. молекул АТР, гидролиз которых до ADP и фосфата обеспечивает необходимой энергией множество энергетически невыгодных реакций.
2.3.2. АТР в процессе гликолиза может образовываться даже в отсутствие кислорода
Самым важным этапом стадии 2 катаболизма является гликолиз последовательность реакций, приводящих к расщеплению глюкозы. При гликолизе молекула глюкозы, содержащая шесть атомов углерода, превращается в две молекулы пирувата, содержащие по три атома углерода каждая. Для такого превращения требуется девять последовательных ферментативных реакций, в которых происходит образование ряда промежуточных фосфатсодержащих соединений (рис. 2-20). Логически рассуждая, последовательность реакций гликолиза можно разделить на три этапа: 1) в реакциях 1-4 глюкоза превращается в трехуглеродный альдегид глицеральдегид-3-фосфат (для этого превращения нужны две фосфатные группы, а необходимая энергия выделяется при гидролизе АТР); 2) в реакциях 5 и 6 альдегидная группа каждой молекулы глицеральдегид-3-фосфата окисляется до карбоксильной, и выделяющаяся при этом энергия расходуется на синтез АТР из ADP и неорганического фосфата; 3) в реакциях 7, 8 и 9 те две молекулы фосфата, которые присоединились к сахару на первой стадии, переносятся обратно на ADP, в результате чего образуется АТР и компенсируются затраты АТР на этапе 1.
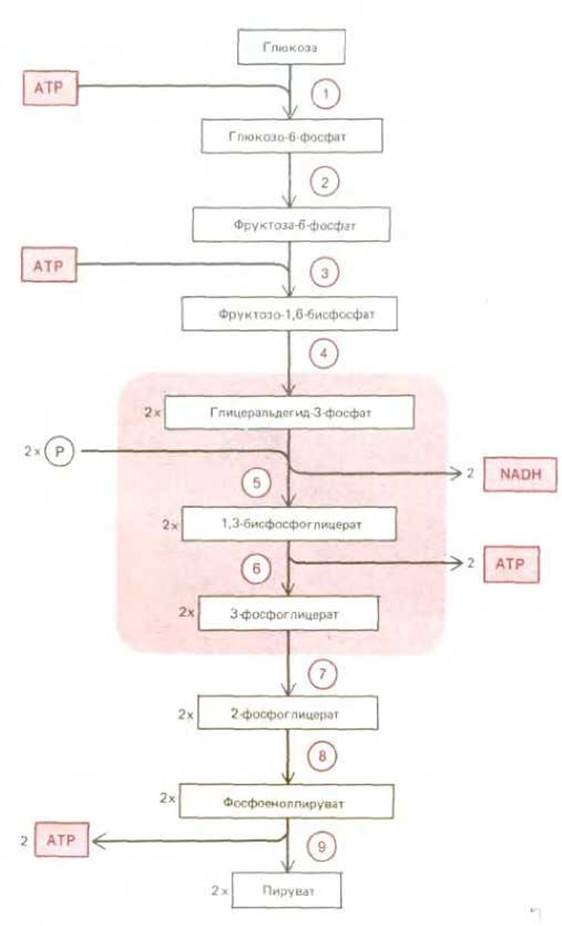
Рис. 2-20. Промежуточные продукты гликолиза. Каждая из пронумерованных реакций катализируется особым ферментом. На этапе 4 шестиуглеродный сахар расщепляется, давая два трехуглеродных сахара, так что после этой реакции число молекул на каждом этапе удваивается. Реакции 5 и 6 ответственны за суммарный синтез АТР и NADH (см. рис. 2-21).
Суммарный выход энергии при гликолизе сводится к синтезу двух молекул АТР (на одну молекулу глюкозы), которые образовались в реакциях 5 и 6. Таким образом, данные реакции имеют решающее значение для гликолиза. Эти две реакции - единственные во всем процессе, в которых из неорганического фосфата формируется высокоэнергетическая фосфатная связь, - иллюстрируют, каким образом может осуществляться взаимосвязь внутриклеточных реакций для реализации выделяющейся при окислении энергии (рис. 2-21). Суммарным результатом этих двух реакций является окисление сахарного альдегида в фосфоглицероловую кислоту, перенос неорганического фосфата на ADP с образованием высокоэнергетической связи АТР и восстановление NAD+ до NADH (рис. 2-22). Возможно, что в процессе эволюции эта изящная пара сопряженных реакций возникла на самых ранних этапах метаболизма. Кроме того, имея первостепенное значение для метаболизма глюкозы, эти реакции в процессе фотосинтеза протекают в противоположном направлении за счет NADPH и АТР, образующихся в реакциях, активируемых светом. В связи с этим они играют центральную роль в фотосинтетическом процессе фиксации углерода на стадии образования глицеральдегид-3-фосфата (см. разд. 7.3.4).
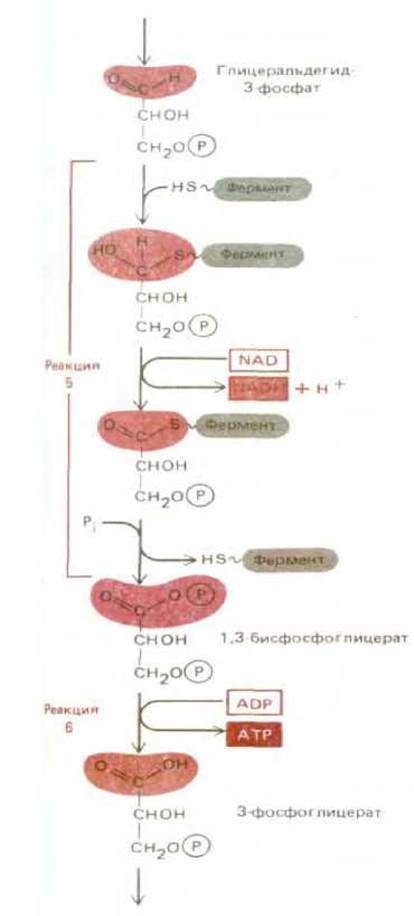
Рис. 2-21. Этапы 5 и 6 гликолиза: окисление альдегида до карбоновой кислоты сопряжено с образованием АТР и NADH (см. также рис. 2-20). Этап 5 начинается с образования ковалентной связи между ферментом глицеральдегид-3-фосфатдегидрогеназой и углеродом альдегидной группы глицеральдегид-3-фосфата. Затем углерод (в виде гидрид-иона: протона плюс двух электронов) отщепляется от связанной с ферментом альдегидной группы глицеральдегид-3-фосфата и переносится к молекуле-переносчику NAD+ (см. рис. 2-22). В результате этой окислительной стадии образуется карбонильная группа сахара, соединенная с ферментом высокоэнергетической связью. Затем фосфат-ион из раствора разрывает эту связь, образуя вместо нее высокоэнергетическую сахарофосфатную связь. В двух этих последних реакциях фермент обеспечивает сопряжение энергетически выгодного процесса окисления альдегида с энергетически невыгодным процессом образования высокоэнергетической связи. Наконец, на этапе 6 гликолиза новообразованная реактивная фосфатная группа переносится на ADP, вследствие чего образуется АТР, а на окисленном сахаре остается свободная карбоксильная группа.
Для большинства клеток животных гликолиз служит лишь прелюдией к стадии 3 катаболизма, так как образующаяся при гликолизе молочная кислота быстро поступает в митохондрии, где полностью окисляется до СО2 и Н2О. Тем не менее у анаэробных организмов (т. е. таких, которые не используют молекулярного кислорода) и тканей (например, скелетных мышц), способных работать в анаэробных условиях, гликолиз может стать основным источником клеточного АТР. В этих случаях вместо того, чтобы подвергнуться расщеплению в митохондриях, молекулы пирувата остаются в цитозоле и в зависимости от вида организма могут превращаться либо в этанол плюс СО2 (в дрожжах), либо в лактат (в мышцах), которые затем выводятся из клеток. Дальнейшее превращение пирувата в этих энергодающих реакциях, называемых брожением, требуется для того, чтобы полностью использовать восстановительный потенциал, полученный в реакции 5 гликолиза, и таким путем регенерировать NAD+, необходимый для дальнейшего осуществления гликолиза (см. разд. 7.4.1).
2.3.3. Окислительный катаболизм поставляет значительно большее количество биологически полезной энергии [14]
Анаэробное образование АТР из глюкозы в реакциях гликолиза относительно неэффективно. Конечные продукты анаэробного гликолиза все еще несут в себе очень большое количество химической энергии, которая может выделиться при последующем окислении. Развитие окислительного катаболизма (клеточное дыхание) стало возможным лишь после того, как в результате фотосинтеза, осуществляемого цианобактериями, в атмосфере Земли было накоплено достаточное количество молекулярного кислорода. До тех пор, пока это не произошло, доминирующей формой поддержания жизни были, по-видимому, анаэробные катаболические процессы. Добавление к катаболическому процессу стадии, требующей присутствия кислорода (стадия 3 на рис. 2-18), обеспечивает клетки значительно более мощным и эффективным методом извлечения энергии из молекул питательных веществ. Эта стадия (стадия 3) начинается с цикла лимонной кислоты (его называют также циклом трикарбоновых кислот или циклом Кребса) и завершается окислительным фосфорилированием: оба процесса имеют место в аэробных бактериях и митохондриях эукариотических клеток.
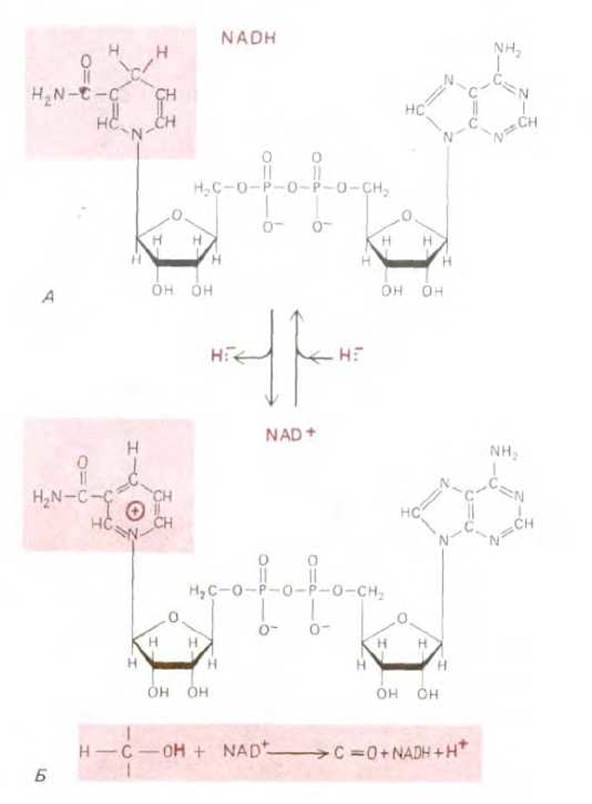
Рис. 2-22. NADH и NAD+ - наиболее важные переносчики водорода в катаболических реакциях. А. Строение молекул NADH и NAD+. NAD это сокращенное обозначение никотинамидадениндинуклеотида; в самом названии отражена структура молекулы, правая часть (на рисунке) которой представлена аденозинмонофосфатом (AMP). Часть молекулы NAD+, называемая никотинамидным кольцом (в цветном прямоугольнике), может присоединять атом водорода с лишним электроном (гидрид-ион, Н~), восстанавливаясь до NADH. В такой восстановленной форме никотинамидное кольцо менее стабильно, поскольку в этом случае отсутствует стабилизирующее влияние резонанса. В результате присоединившийся гидрид-ион переносится на другие молекулы. Б. Пример реакции с участием NAD+ и NADH. При биологическом окислении молекулы субстрата, например спирта, субстрат теряет два атома водорода. Один из них в виде гидрид-иона присоединяется к NAD+, образуя NADH, тогда как другой выделяется в раствор в виде протона Н+ (см. рис. 7-18).
2.3.4. Центральным процессом метаболизма является цикл лимонной кислоты [15]
Главная функция цикла лимонной кислоты - окисление ацетогруппы, включающейся в этот цикл в форме молекул ацетил-СоА. Процесс этот носит циклический характер, поскольку ацетогруппа окисляется не сразу, а лишь после того, как она ковалентно присоединится к более крупной молекуле - оксалоаиетату, которая регенерируется после каждого оборота цикла. Как показано на рис. 2-23, цикл начинается с реакции ацетил-СоА с оксалоацетатом, приводящей к образованию молекулы трикарбоновой кислоты, называемой лимонной кислотой (или цитратом). Затем следует серия реакций, в которых два из шести атомов углерода цитрата окисляются до СО2, образуя молекулу оксалоацетата - исходного продукта для нового цикла. (Поскольку два новых атома углерода, присоединяемых в каждом цикле, входят не в ту часть молекулы цитрата, которая окисляется в данном цикле до СО2, должно пройти несколько циклов, прежде чем подойдет их очередь окислиться.) Молекулы СО2, образующиеся в подобных реакциях, затем диффундируют из митохондрий (или из бактерий) и покидают клетку.
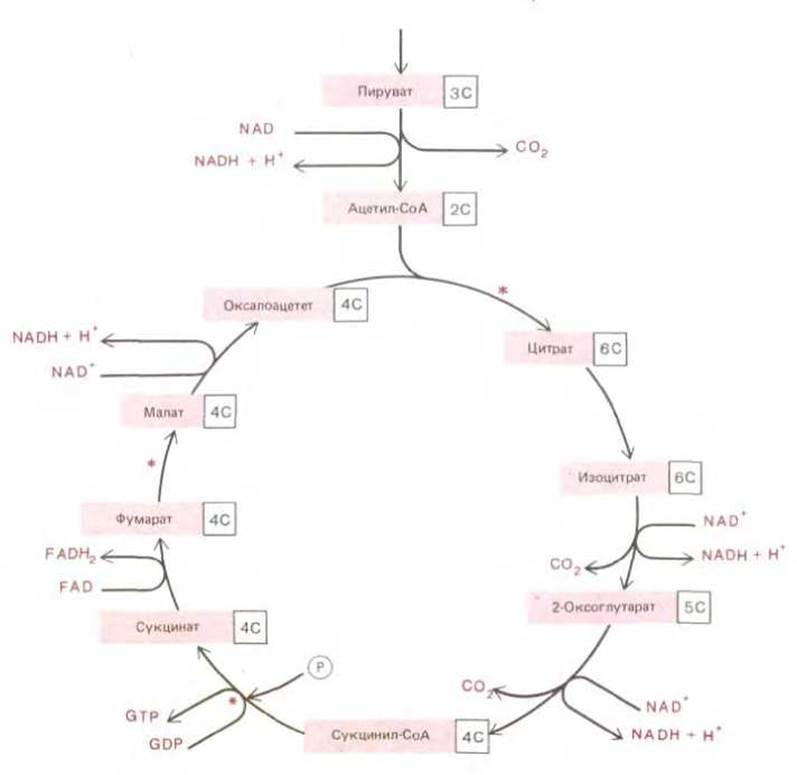
Рис. 2-23. Цикл лимонной кислоты. В митохондриях и клетках аэробных бактерий ацетогруппы, образованные из пирувата, подвергаются дальнейшему окислению. Атом углерода ацетильной группы превращается в СО2, водородные же атомы переносятся к молекулам-переносчикам NAD+ и FAD. Дополнительные атомы кислорода и водорода включаются в цикл в виде молекул воды на стадиях, отмеченных звездочками (*). Более детально цикл лимонной кислоты показан на рис. 7-14.
Энергия, высвобождающаяся при окислении связей С—Н и С—С цитрата, потребляется несколькими различными способами в цикле лимонной кислоты. В одной из реакций цикла (сукцинил-СоА — сукцинат) высокоэнергетическая фосфатная связь образуется под действием механизма, сходного с тем, который мы уже рассмотрели в случае гликолиза. (Хотя в реакции цикла образуется не АТР, a GTP, все нуклеозидтрифосфаты равноценны в энергетическом отношении благодаря реакциям обмена типа ADP + GTR ⇄ ATP + GDP.) Оставшаяся часть энергии, полученной при окислении, расходуется на перевод молекул-переносчиков водорода (или гидрид-ионов) в восстановленную форму; в каждом обороте цикла три молекулы NAD+ превращаются в NADH, а одна молекула флавинадениндинуклеотида (FAD) - в FADH2. Энергия, переносимая на таких молекулах- переносчиках активированными атомами водорода, используется в реакциях окислительного фосфорилирования; для осуществления последних (они будут подробно рассмотрены ниже) необходим молекулярный кислород атмосферы.
Дополнительные атомы кислорода, необходимые для образования СО2 из включающейся в цикл лимонной кислоты ацетильной группы, поставляются не молекулярным кислородом, а молекулой воды. В каждом цикле расщепляются три молекулы воды, атомы кислорода которых используются для образования СО2. Некоторые из атомов водорода молекул воды связываются с молекулами субстрата, переходя в более высокое энергетическое состояние, и затем переносятся (вместе с атомами водорода ацетогруппы) к таким молекулам-переносчикам, как NADH.
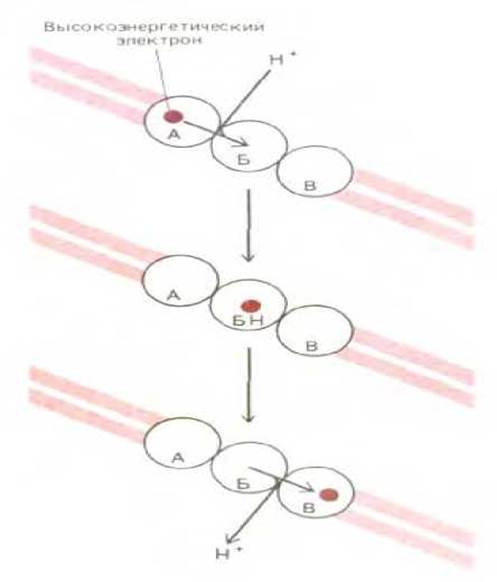
Рис. 2-24. Возникновение трансмембранного протонного градиента в результате реакций переноса электронов. Высокоэнергетический электрон (полученный, например, при окислении метаболита) последовательно переводится переносчиками А, Б и В в более низкое энергетическое состояние. На данной схеме переносчик Б располагается в мембране таким образом, что при прохождении электрона он захватывает ион Н+ с одной стороны от мембраны и высвобождает его с другой. Возникающий в результате градиент Н+ представляет собой форму запасания энергии; эта энергия используется другими белками митохондриальной мембраны для инициирования синтеза АТР (см. рис. 7-35).
В эукариотических клетках митохондрия - это центр, к которому ведут все катаболические пути независимо от того, что служит для них первоначальным субстратом-сахара, жиры или белки. Объясняется это тем, что не только пируват, но и жирные кислоты, равно как и некоторые аминокислоты, тоже поступают из цитозоля в митохондрии, где они превращаются в ацетил-СоА или в один из промежуточных продуктов цикла лимонной кислоты. Митохондрия служит также отправной точкой биосинтетических реакций, поскольку в ней образуются такие жизненно важные углеродсодержащие промежуточные продукты, как оксалоацетат и 2-оксоглутарат. Эти соединения переносятся из митохондрии обратно в цитозоль, где они используются в качестве предшественников таких важнейших молекул клетки, как, например, аминокислоты.
2.3.5. При окислительном фосфорилировании перенос электронов к кислороду приводит к образованию АТР [9, 16]
Окислительное фосфорилирование является последней стадией катаболизма; в ходе этого процесса высвобождается большая часть метаболической энергии. При окислительном фосфорилировании молекулы NADH и FADH2 переносят электроны, полученные ими от кислорода молекул питательных веществ, к молекулярному кислороду О2. В этой реакции, которая формально равноценна сгоранию водорода в воздухе с образованием воды, высвобождается значительное количество химической энергии. Часть этой энергии используется для образования АТР; остальная энергия выделяется в виде тепла.
Хотя при окислении NADH и FADH2 в конечном счете происходит перенос водорода к кислороду, водород транспортируется не в атомарном состоянии. Объектом переноса в данном случае служат электроны атома водорода. Это связано с тем, что атом водорода может легко диссоциировать на свои составные части - электрон и протон (Н+). Затем электрон может отдельно переноситься к молекуле, акцептирующей лишь электроны, а протоны при этом остаются в водном растворе. По той же причине в случае, когда к молекуле с сильным сродством к водороду присоединяется лишь электрон, автоматически образуется атом водорода, так как из раствора сразу же акцептируется протон. В ходе окислительного фосфорилирования электроны от NADH и FADH2 переносятся по цепи молекул-переносчиков, но при этом форма, в которой они транспортируются (молекула водорода или электрон), зависит от природы переносчика.
В эукариотической клетке эта последовательность реакций переноса электронов по электрон-транспортной цепи локализована на внутренней митохондриальной мембране, в которую встроены все молекулы-переносчики. Перенос электронов сопровождается уменьшением их энергии на каждой ступени вплоть до конечной точки процесса, где электроны переносятся на молекулы кислорода. Поскольку молекулы кислорода обладают наибольшим сродством к электронам, связываясь с кислородом, электроны оказываются на самом нижнем энергетическом уровне. Энергия, выделяющаяся при переходе электронов на более низкие энергетические уровни, используется для перекачивания протонов из внутреннего пространства митохондрии наружу, причем механизм этого
процесса до конца еще не выяснен (рис. 2-24). В результате во внутренней митохондриальной мембране создается трансмембранный электрохимический градиент протонов. Этот градиент в свою очередь обусловливает обратное перемещение протонов через ферментный комплекс в мембране; особый фермент этого комплекса (АТР-синтетаза) катализирует присоединение фосфатной группы к ADP с образованием в митохондрии АТР. Вновь синтезированный АТР переносится из митохондрии в другие части клетки, где он используется для осуществления множества метаболических реакций.
Природа электрон-транспортной цепи и механизм синтеза АТР подробно рассматриваются в гл. 7.
2.3.6. Аминокислоты и нуклеотиды принимают участие в круговороте азота
В метаболических процессах, рассмотренных в предыдущих разделах, участвовали главным образом углеводороды. До сих пор мы не обсуждали метаболизма азота или серы. Эти два элемента являются важными компонентами белков и нуклеиновых кислот - двух самых необходимых макромолекул клетки, на долю которых приходится 2/3 ее сухой массы. Претерпевая различные превращения в ряде обратимых циклических процессов, атомы азота и серы переходят из одного соединения в другое и из тканей различных организмов во внешнюю среду.
Молекулярный азот в изобилии содержится в земной атмосфере, однако химически он неактивен. Лишь небольшое число видов живых существ способно непосредственно включать азот в органические молекулы с помощью процесса, называемого азотфиксацией. Фиксация азота осуществляется особыми микроорганизмами. Кроме того, она происходит и при некоторых геофизических процессах, таких, как вспышка молнии. Фиксация азота имеет критическое значение для биосферы, так как без этого процесса жизнь на нашей планете была бы невозможна. Тем не менее у современных организмов лишь небольшая часть азотистых соединений образуется в результате непосредственной фиксации азота. Основная масса органического азота циркулирует в течение некоторого времени, переходя от одного организма к другому. Таким образом, можно сказать, что азотфиксирующие реакции обеспечивают пополнение общих запасов азота.
Позвоночные получают практически весь азот из содержащихся в их рационе белков и нуклеиновых кислот. В организме эти макромолекулы расщепляются до аминокислот или нуклеотидов, из которых потом образуются новые белки и нуклеиновые кислоты или другие молекулы. Около половины из 20 аминокислот, входящих в состав белков, представляют собой незаменимые аминокислоты (рис. 2-25): они не могут синтезироваться в организме и должны обязательно поступать с пищей. Остальные аминокислоты могут синтезироваться с использованием разнообразных исходных веществ, в число которых входят и промежуточные продукты цикла лимонной кислоты. Незаменимые аминокислоты образуются в организмах других живых существ - обычно в ходе длительных и энергоемких процессов; метаболические пути этих процессов были утеряны позвоночными в ходе эволюции.
Нуклеотиды, необходимые для образования ДНК и РНК, могут синтезироваться в ходе специализированных биосинтетических процессов: не существует «незаменимых нуклеотидов», которые должны непременно поступать в организм в составе пищи. Все атомы азота, входящие в состав пуриновых и пиримидиновых оснований (как и некоторые углеродные атомы), попадают в них из богатых азотом аминокислот - глутамина, аспарагиновой кислоты и глицина, а рибоза и дезоксирибоза являются производными глюкозы.

Рис. 2-25. Девять незаменимых аминокислот, которые не синтезируются в клетках человека и поэтому должны поступать в организм с пищей.
Аминокислоты, не используемые для биосинтеза, могут окисляться, выделяя метаболическую энергию. Большинство входящих в них атомов углерода и водорода образуют в итоге СО2 и Н2О. Атомы же азота, многократно переходя из одной формы в другую, в конце концов выводятся из организма в виде мочевины. Пути превращений различных аминокислот не одинаковы, и метаболизм этих соединений включает множество разнообразных ферментативных реакций.
Заключение
Считается, что клетки животных извлекают энергию из пищи в три этапа. На первом этапе белки, полисахариды и жиры расщепляются в результате внеклеточных реакций на малые молекулы. На втором этапе эти малые молекулы расщепляются в клетках с образованием ацетил-СоА, а также небольшого количества АТР и NADH. Такие реакции - единственные, в которых энергия может выделяться и в отсутствие кислорода. На третьей стадии молекулы ацетил-СоА расщепляются в митохондриях, образуя СО2 и атомы водорода, которые связываются с молекулами таких переносчиков, как NADH. Электроны от атомов водорода переходят по сложной цепи переносчиков, что в конечном счете приводит к восстановлению молекулярного кислорода и образованию воды. Под действием энергии, высвобождающийся на разных стадиях переноса электронов, ионы водорода (Н+) транспортируются из внутреннего пространства митохондрии наружу. Возникающий в результате трансмембранный электрохимический градиент протонов во внутренней митохондриальной мембране поставляет энергию для синтеза основного количества молекул АТР клетки.