Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Поддержание нормальной организации тканей
Обновление за счет стволовых клеток. Пример: эпидермис
Перейдем теперь от клеточных популяций, обновляющихся путем простого удвоения своих клеток, к таким, которые обновляются за счет стволовых клеток. Эти популяции сильно различаются не только по свойствам самих клеток и скорости их замещения, но и по пространственной организации этого процесса. Например, в выстилке тонкого кишечника клетки образуют однослойный эпителий. Этот эпителий покрывает поверхность ворсинок, выступающих в просвет кишки, и он же выстилает глубокие крипты, уходящие в толщу подлежащей соединительной ткани (рис. 17-17). Стволовые клетки находятся в защищенном месте в глубине крипт. Дифференцированные клетки, образующиеся из стволовых, выносятся в результате скольжения их в плоскости эпителиального слоя наверх, пока не достигнут открытой поверхности ворсинок, с кончиков которых они в конце концов слущиваются. Примером совсем иного процесса может служить кожа: эпидермис представляет собой многослойный эпителий, и дифференцирующиеся клетки перемещаются от места их образования в направлении, перпендикулярном плоскости клеточных слоев. В кроветворных тканях пространственная картина образования клеток сложна и выглядит хаотичной. Но прежде чем углубляться в дальнейшие подробности, посмотрим, что представляет собой стволовая клетка.
17.4.1. Стволовые клетки обладают способностью неограниченно делиться и давать дифференцированное потомство [17]
Для стволовой клетки определяющим будут следующие свойства:
1) она сама не является терминально дифференцированной (т. е. не прошла путь дифференцировки до конца);
2) она способна к неограниченному делению (по крайней мере в течение жизни организма);
3) при ее делении каждая дочерняя клетка стоит перед выбором - остаться стволовой клеткой, какой была родительская, или встать на путь, необратимо ведущий к полной дифференцировке (рис. 17-18).
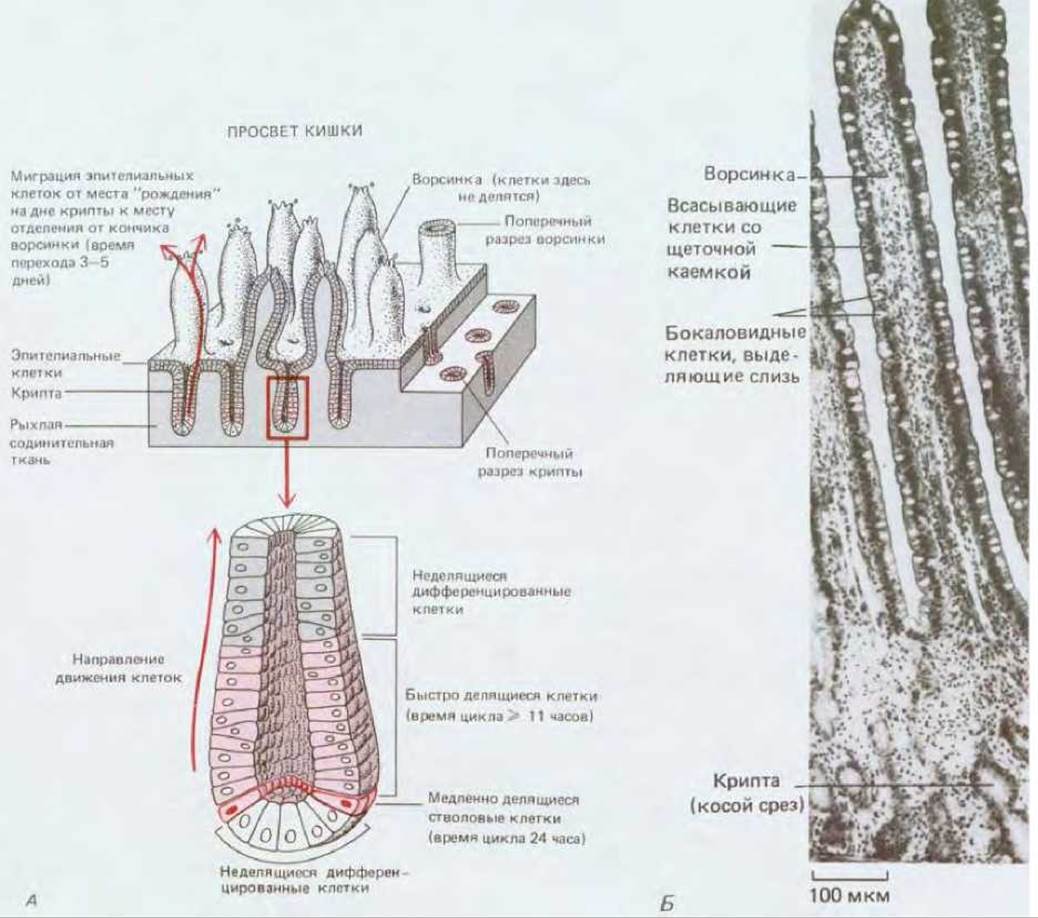
Рис. 17-17. А. Схема обновления клеточной популяции в выстилке тонкой кишки за счет пролиферации стволовых клеток. Б. Участок среза, на котором можно видеть ворсинки и крипты. Обратите внимание на светлые бокаловидные клетки (они выделяют слизь), разбросанные в эпителии ворсинок среди всасывающих клеток со щеточной каемкой. См. также рис. 17-9, где показана структура этих клеток. (С любезного разрешения Peter Gould.)
Стволовые клетки нужны в любом месте, где постоянно возникает потребность в новых дифференцированных клетках, которые, однако, сами делиться не могут. В ряде тканей конечное состояние дифференцировки явно несовместимо с клеточным делением. Например, ядра клеток могут разрушаться, как это происходит в наружных слоях эпидермиса, или выталкиваться из клеток, как при созревании эритроцитов у млекопитающих. Иногда осуществлению митоза и цитокинеза препятствует то, что цитоплазма плотно заполнена таким материалом, как, например, миофибриллы мышечных клеток. В других терминально дифференцированных клетках невозможность деления может быть обусловлена какими-то более тонкими биохимическими причинами. В любом таком случае обновление будет зависеть от стволовых клеток.
Стволовые клетки предназначены не для того, чтобы выполнять определенную специализированную функцию, а для того, чтобы производить клетки с такой функцией. Поэтому стволовым клеткам часто не свойствен какой-либо характерный вид, и тогда их трудно опознать. Но это не означает, что все они одинаковы. Не будучи внешне дифференцированы, они тем не менее детерминированы (разд. 16.2.8): клетка-сателлит в скелетной мышце - как источник мышечных волокон, базальная клетка эпидермиса - как источник ороговевающих эпителиальных клеток, сперматогония - как источник спермиев, базальная клетка обонятельного эпителия - как источник обонятельных нейронов (рис. 17-19) и т. д. Стволовые клетки, порождающие только один вид дифференцированных клеток, называют унипотентными, а те, которые порождают несколько их видов - плюрипотентными.
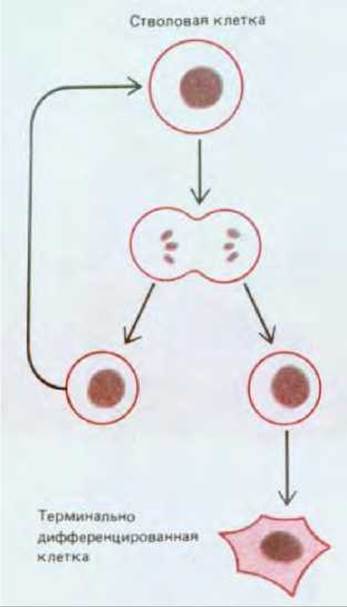
Рис. 17-18. Судьба потомков стволовой клетки. Каждая дочерняя клетка, образующаяся при делении стволовой клетки, может остаться также стволовой, а может пойти по пути, ведущему к терминальной дифференцировке.
При изучении тканей, образующихся из стволовых клеток, возникает много серьезных вопросов. Чем определяется, будет ли данная стволовая клетка делиться или останется в состоянии покоя? От чего зависит, станет ли дочерняя клетка тоже стволовой или начнет дифференцироваться? И если она вступит на путь дифференцировки, то чем этот процесс будет регулироваться? Мы начнем рассмотрение с эпидермиса, так как его простая пространственная организация облегчает изучение биологии его стволовых клеток и судьбы их потомства.
17.4.2. Эпидермис подразделен на пролиферативные единицы [18, 19]
Эпидермальный слой кожи и эпителиальная выстилка пищеварительного тракта - это две ткани, наиболее подверженные прямым повреждающим воздействиям со стороны внешнего мира. В обеих тканях зрелые дифференцированные клетки быстро снашиваются в самых уязвимых участках и так же быстро замещаются в результате пролиферации менее дифференцированных клеток, находящихся в более защищенных местах.
Эпидермис - это многослойный эпителий, состоящий в основном из кератиноцитов (называемых так потому, что характерной чертой их дифференцированного состояния является синтез кератина) (рис. 17-20). От слоя к слою эти клетки изменяют свой внешний вид. Самый глубокий из внутренних слоев образован базальными клетками. В основном именно эти клетки делятся путем митоза. Над базальными клетками находится несколько слоев более крупных шиповатых клеток (рис. 17-21). Такое название они получили благодаря своему виду на препаратах для световой микроскопии: их многочисленные десмосомы с отходящими от них толстыми пучками кератиновых волокон едва различимы и выглядят как крошечные шипы на поверхности клеток. Выше шиповатых клеток лежит тонкий слой зернистых клеток (рис. 17-20); он образует границу между внутренней метаболически активной зоной и самым наружным слоем, состоящим из мертвых клеток, в которых все внутриклеточные органеллы исчезли. Эти наружные клетки редуцированы до плоских чешуек, заполненных плотно упакованным кератином. На поверхности, обращенной к цитоплазме, плазматические мембраны чешуек и наружных зернистых клеток укреплены тонким (12 нм), жестким слоем с поперечными сшивками, содержащим внутриклеточный белок инволюкрин. Сами чешуйки обычно настолько уплощены, что границы их в световом микроскопе почти неразличимы; но если выдержать препарат в растворе NaOH, эти клетки несколько набухнут, и тогда после надлежащей окраски можно увидеть (если эпидермис в данном участке тонкий) удивительно правильную геометрическую картину расположения клеток: чешуйки уложены здесь гексагональными колонками, которые аккуратно сцеплены между собой краями клеток (рис. 17-22). Ширина колонок такова, что под каждой из них, в ее основании, находится около десятка базальных клеток. Эти клетки можно подразделить на центральные и периферические в соответствии с их положением в основании колонки. Периферические (но не центральные!) клетки иногда можно видеть в тот момент, когда они переходят из базального слоя наверх, в слой шиповатых клеток. Каждую колонку называют пролиферативной единицей эпидермиса. Хотя упорядоченная организация в виде правильных колонок выявляется только в некоторых участках кожи, она служит хорошей иллюстрацией общих принципов обновления эпидермальных клеток.
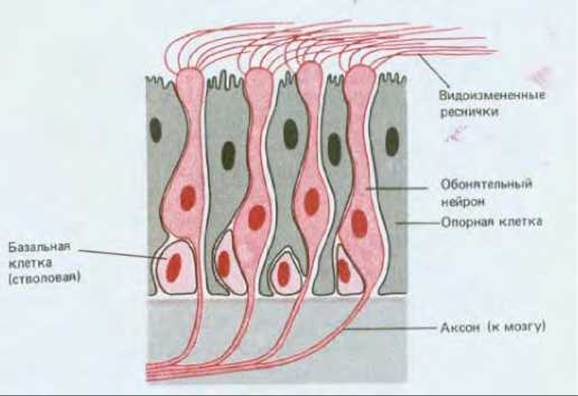
Рис. 17-19. Схема строения обонятельного эпителия, специализированного для восприятия запахов. Здесь можно различить три типа клеток: опорные клетки, базальные клетки и обонятельный нейроны. Как показывают эксперименты с применением радиоавтографии, базальные клетки являются стволовыми: из них образуются обонятельные нейроны. Это редкое исключение из правила, согласно которому нейроны представляют собой перманентные клетки. Каждый обонятельный нейрон служит около месяца (у млекопитающих), а затем заменяется новым. От круглой «головки» обонятельного нейрона отходят 6- 8 видоизмененных ресничек; как полагают, в них-то и содержатся рецепторы для пахучих веществ. Аксон, идущий от другого конца нейрона, передает информацию в мозг. Всякий раз, когда базальная клетка дифференцируется в обонятельный нейрон, от него отрастает новый аксон, который образует надлежащие связи в мозгу.
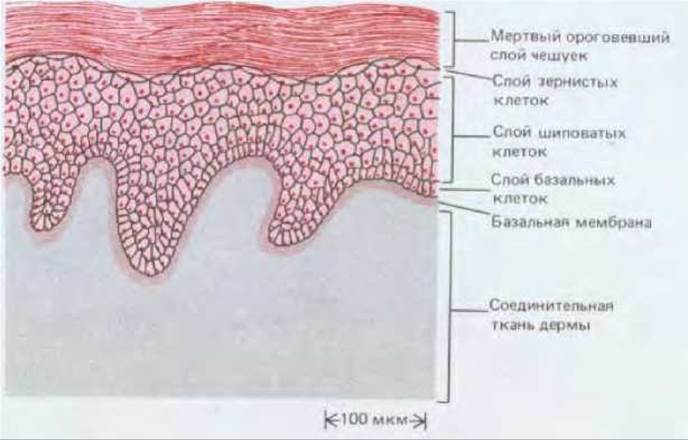
Рис. 17-20. Строение эпидермиса средней толщины у млекопитающего (схема) (см. также рис. 17-1). Зернистые (гранулярные) клетки находятся между шиповатыми клетками и уплощенными чешуйками. Они проходят предпоследнюю стадию ороговения и содержат интенсивно окрашивающиеся гранулы малоизученного материала - кератогиалина, участвующего в уплотнении и перекрестном сшивании кератина внутри клетки. Кератогиалин в основном состоит из белка филаггрина. Кроме клеток, которым предстоит ороговение, в глубоких слоях эпидермиса находится небольшое число клеток совсем иного типа (на схеме не показанных): это макрофагоподобные клетки Лангерганса, происходящие из костного мозга; меланоциты, происходящие из нервного гребня; клетки Меркеля, связанные с нервными окончаниями в эпидермисе.
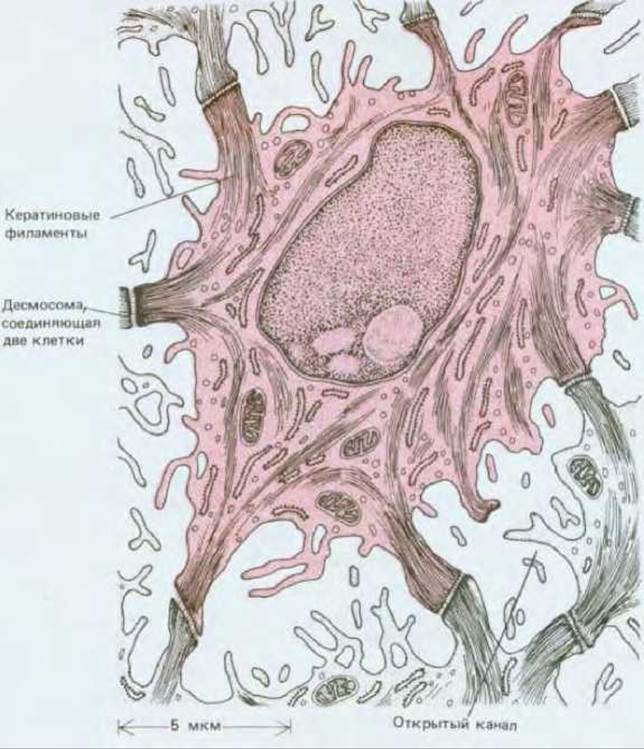
Рис. 17-21. Рисунок, сделанный по электронной микрофотографии среза шиповатой клетки эпидермиса (выделена цветом). Видны пучки кератиновых нитей, которые пронизывают цитоплазму и направляются к десмосомам, соединяющим клетку с ее соседями. Обратите внимание, что между соседними клетками есть открытые каналы, позволяющие питательным веществам свободно диффундировать через метаболически активные слои зпидермиса. Ближе к его поверхности, на уровне зернистых клеток, имеется водонепроницаемый барьер, образованный, по-видимому, изолирующим веществом, который эти клетки выделяют из особых пузырьков. (R. V. Krstic, Ultrastruture of the Mammalian Cell: An Mas. Berlin: Springer, 1979.)
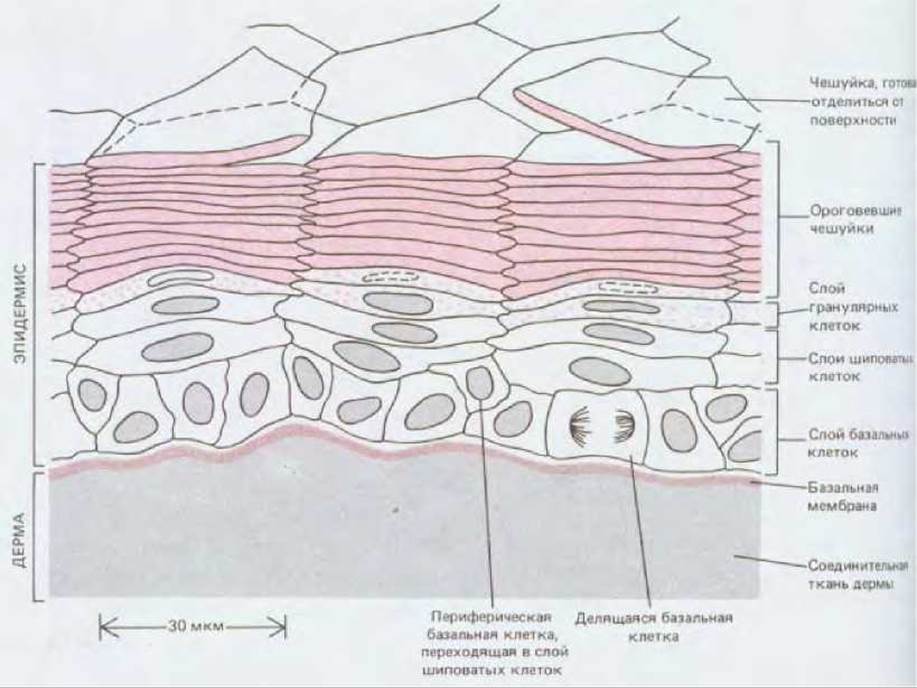
Рис. 17-22. Пролиферативные единицы, или колонки, в эпидермисе тонкой кожи. Эта структура выявляется при набухании ороговевших чешуек в растворе, содержащем NaOH. Такая организация в виде колонок свойственна лишь тонким участкам эпидермиса.
17.4.3. В дифференцирующихся эпидермальных клетках по мере их созревания последовательно синтезируются различные кератины [19]
Перейдем от описанной статической картины к динамике. Центральная базальная клетка колонки делится, и некоторые из дочерних клеток, в свою очередь поделившись, сдвигаются к периферии основания. Периферические базальные клетки переходят из базального слоя в слой шиповатых клеток - на первую ступень движущегося вверх «эскалатора». Достигнув зернистого слоя, шиповатые клетки начинают терять свои ядра и цитоплазматические органеллы и постепенно превращаются в ороговевшие чешуйки наружного слоя. В конце концов эти чешуйки отслаиваются и разносятся токами воздуха, образуя один из главных компонентов комнатной пыли. У человека промежуток времени от момента рождения клетки в базальном слое эпидермиса до ее слущивания с поверхности кожи занимает от двух до четырех недель в зависимости от участка тела.
Сопутствующие химические изменения можно изучать, анализируя тонкие слои эпидермиса, срезанные параллельно поверхности, или последовательные слои клеток, обдираемые при повторном наложении и снятии кусков липкой ленты. При этом можно экстрагировать и охарактеризовать молекулы кератина, которых очень много во всех слоях эпидермиса. Существует множество различных видов кератина (разд. 11.5.1), кодируемых большим семейством гомологичных генов: благодаря меняющемуся прецессингу их транскриптов разнообразие кератинов еще больше возрастает. По мере того как стволовая клетка. находившаяся в основании колонки, превращается в чешуйку наверху (рис. 17-22), в ней последовательно экспрессируются различные выборки из всего комплекта гомологичных кератиновых генов. В ходе этого процесса начинают синтезироваться другие характерные белки, такие как инволюкрин, и это тоже входит в координированную программу терминальной дифференцировки клеток.

Рис. 17-23. Каждая пролиферативная единица должна всегда содержать по меньшей мере одну «бессмертную» стволовую клетку, потомки которой будут находиться в этой единице и в отдаленном будущем. Стрелками показано происхождение одних клеток от других. Стволовая клетка в каждой клеточной генерации представлена здесь в центральном положении. Другие базальные клетки могут изначально обладать иными химическими свойствами, которые предопределяют их уход из базального слоя и дифференцировку; в другом варианте базальные клетки будут эквивалентны бессмертным стволовым клеткам по своим свойствам, но их потомство может выталкиваться из базального слоя и слущиваться с кожи, и в этом смысле такие базальные клетки будут смертными.
17.4.4. Возможно, что «бессмертие» стволовой клетки сохраняется благодаря контакту с базальной мембраной [20]
Если каждая пролиферативная единица эпидермиса поддерживается неопределенно долго за счет размножения ее базальных клеток, то среди них должна быть хотя бы одна клетка, потомство которой не вымирает полностью до конца жизни животного. Мы будем называть такую клетку бессмертной стволовой клеткой (рис. 17-23). В принципе деление бессмертной стволовой клетки могло бы давать две первоначально одинаковые дочерние клетки, чья дальнейшая судьба зависела бы уже от последующих условий их жизни. В противоположном крайнем случае деление стволовой клетки могло бы всегда быть асимметричным, так что одна и только одна из дочерних клеток наследовала бы свойства, необходимые для бессмертия; в другой же клетке что-то изменялось бы уже в момент ее образования, и это заставляло бы ее дифференцироваться и обрекало в конце концов на гибель. В таком случае число бессмертных стволовых клеток никогда не могло бы увеличиться, а это противоречит фактам. Если участок эпидермиса разрушен, непрерывность ткани восстанавливают окружающие здоровые эпидермальные клетки, которые мигрируют и размножаются, чтобы закрыть брешь. При этом образуются новые пролиферативные единицы, и их центральные базальные клетки неизбежно должны были возникнуть в результате таких делений, когда из одной бессмертной клетки получаются две.
Таким образом, при делении стволовой клетки судьба дочерних клеток должна хотя бы отчасти зависеть от внешних факторов. Одним из таких факторов мог бы быть контакт с базальной мембраной, разрыв которого приводил бы к запуску терминальной дифференцировки. Эксперименты на тканевых культурах в какой-то мере подтверждают это предположение: эпидермальные клетки продолжают делиться, если они растут в контакте с подходящим субстратом (например, со слоем фибробластов), но сразу начинают дифференцироваться при росте в суспензии. Однако такого рода регуляция внешними факторами не позволяет объяснить все. Другие данные указывают скорее на обратное направление причинно-следственной связи, т. е. на то, что изменения, ведущие к терминальной дифференцировке, приводят к откреплению клеток от базальной мембраны, а не наоборот. Согласно этой гипотезе, лишь немногие из базальных клеток способны быть стволовыми. Поверхность этих стволовых клеток имеет особые свойства, которые позволяют им прикрепляться к базальной мембране. Эти клетки запрограммированы на то, чтобы давать определенную долю потомства, «обреченную» на дифференцировку, в которую входит и потеря способности к прикреплению. В особых условиях тканевой репарации отношение дифференцирующихся клеток-потомков к пролиферирующим может быть изменено под действием местных ростовых факторов так, чтобы образовались дополнительные клетки для покрытия раны. Данные в пользу этой гипотезы были получены в экспериментах, в которых кератиноциты выращивали in vitro в условиях недостатка кальция; это удерживало их в состоянии монослоя, так что все клетки были базальными. Тем не менее некоторые клетки в этих условиях вступали на путь терминальной дифференцировки, на что указывал синтез инволюкрина; эти дифференцирующиеся клетки выходили из базального слоя, как только повышалась концентрация Са2+ в среде (разд. 14.3.4).
17.4.5. Пролиферация базальных клеток регулируется в соответствии с толщиной эпидермиса [21]
Чем бы ни определялся выбор между сохранением статуса стволовой клетки и переходом на гибельный путь терминальной дифференцировки, должны действовать другие факторы, которые регулировали бы скорость образования новых эпидермальных клеток. Например, если наружные слои эпидермиса соскоблить, то базальные клетки начинают делиться быстрее. Через некоторое время это приводит к восстановлению нормальной толщины эпидермиса, и пролиферация в базальном слое снова снижается до обычного уровня. Все происходит так, как будто удаление наружных дифференцированных слоев освобождает базальные клетки от влияния какого-то ингибирующего фактора, который вновь начинает действовать, как только эпидермис полностью восстанавливается.
Хотя известно, что кератиноциты, растущие в культуре, реагируют на многие гормоны и ростовые факторы, включая фактор роста эпидермиса, очень важный для клиницистов вопрос о молекулярных механизмах, регулирующих пролиферацию этих клеток in vivo, до сих пор не выяснен. Последствия нарушенной регуляции этого процесса можно наблюдать при псориазе. При этом распространенном заболевании кожи пролиферация базальных клеток сильно ускорена, эпидермис утолщен и клетки слущиваются с поверхности кожи уже через неделю после их образования в базальном слое, еще не успев подвергнуться полному ороговению.
17.4.6. Секреторные клетки кожи обособлены в железах, и их популяциям свойственна иная динамика [22]
Кожа служит не только защитным барьером - она выполняет и другие функции. В определенных специализированных участках наряду с описанными выше ороговевшими клетками из эпидермиса развиваются и клетки иных типов. В частности, секреторные клетки обособлены в глубоко лежащих железах, и обновление их происходит совершенно иначе, чем в участках ороговения.
Простейшим примером подобной структуры может служить потовая железа. Она состоит из длинной трубки со слепым концом и образуется как впячивание эпидермиса. Пот выделяют клетки нижней части этой трубки, и он выходит на поверхность кожи через выводной проток (рис. 17-24). Секреторные клетки образуют однослойный эпителий, окруженный небольшим числом сократимых миоэпителиальных клеток (см. рис. 17-25, Б и 17-38, Д). Выводной проток выстлан двуслойным эпителием без миоэпителиальных элементов. Сходным образом устроены и железы, выделяющие слезы, ушную серу, слюну и молоко. По крайней мере в слюнных и молочных железах в протоках имеются стволовые клетки, предназначенные для обновления популяции секреторных клеток.
Молочная железа представляет особый интерес в связи с гормональной регуляцией деления и дифференцировки ее клеток. Образование молока должно начинаться, когда рождается ребенок, и прекращаться, когда ребенка отнимают от груди. Когда молочная железа не функционирует, ее железистая ткань состоит из разветвленных систем выводных протоков, погруженных в соединительную ткань и выстланных в секреторных участках одним слоем сравнительно неактивных эпителиальных клеток. На первом этапе подготовки к интенсивной выработке молока гормоны, циркулирующие в крови в период беременности, стимулируют здесь клеточную пролиферацию; концевые отделы протоков растут и ветвятся, образуя небольшие расширения - альвеолы, содержащие секреторные клетки (рис. 17-25). Секреция молока начинается только при стимуляции этих клеток изменившимся набором гормонов в крови матери после рождения ребенка. Когда кормление ребенка грудью прекращается, секреторные клетки дегенерируют, макрофаги уничтожают их остатки, большая часть альвеол исчезает и железа переходит в состояние покоя.
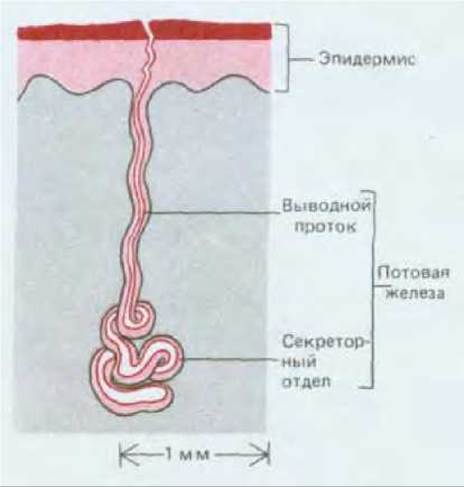
Рис. 17-24.Схема строения потовой железы.
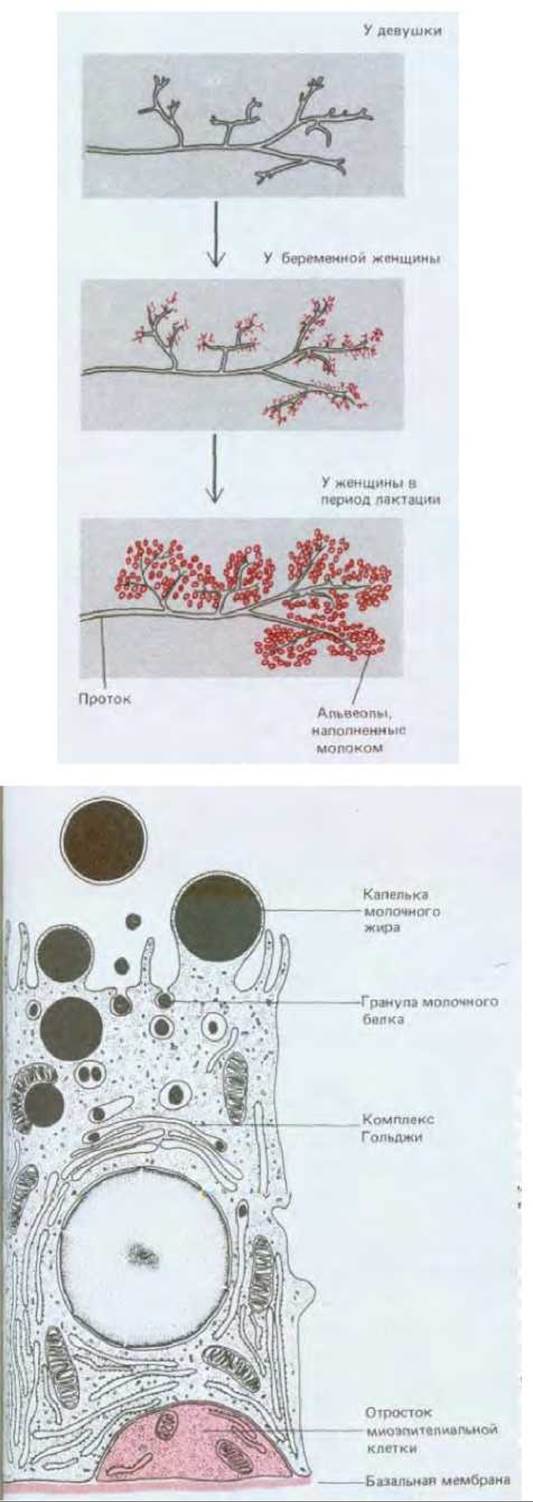
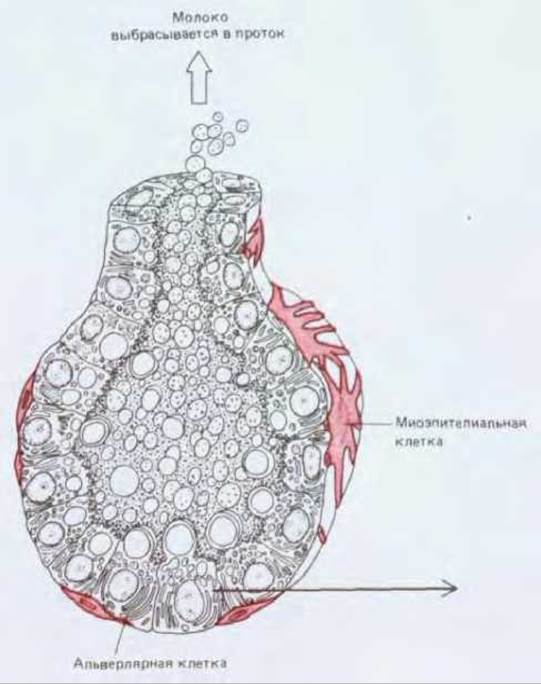
Рис. 17-25. Молочная железа. Вверху слева. Схема образования альвеол из протоков молочной железы во время беременности и лактации. Показан только один небольшой участок железы. В покое железа содержит небольшое количество неактивных железистых элементов, погруженных в массу жировой соединительной ткани (на рисунке - серый фон). Во время беременности происходит сильнейшая пролиферация железистой ткани за счет жировой с преимущественным развитием секреторных отделов железы и образованием альвеол. Вверху справа. Одна из секретирующих молоко альвеол молочной железы и охватывающая ее «корзинка» из миоэпителиальных клеток. Миоэпителиальные клетки сокращаются и выдавливают молоко из альвеол в ответ на воздействие гормона окситоцина, который рефлекторно выделяется у женщины при кормлении грудью. Внизу. Клетки одного и того же типа вырабатывают и молочные белки, и молочный жир. Белки выводятся из клеток путем обычного экзоцитоза, а жир выходит в виде капель, окруженных плазматической мембраной, отделившейся от клетки. (В по R. Krstic, Die Gewebe des Menschen und der Säugetiere. Berlin: Springer-Verlag, 1978; В из D. W. Fawcett, A Textbook of Histology, 11 th ed. Philadelphia: Saunders, 1986.)
Заключение
Многие ткани, особенно те, которым свойственно быстрое замещение клеточной популяции (например, выстилка кишечника, эпидермальный слой кожи, кроветворные ткани), обновляются с помощью стволовых клеток. Стволовые клетки - это, по определению, не до конца дифференцированные клетки, способные неограниченно делиться и давать потомство, часть которого дифференцируется, а часть остается стволовыми клетками. Стволовые клетки эпидермиса лежат в базальном слое, контактируя с базальной мембраной. Потомки стволовых клеток дифференцируются, уходя из этого слоя, и по мере удаления от него последовательно синтезируют различные виды кератинов; затем ядра в клетках дегенерируют, и образуется наружный слой мертвых ороговевших клеток, которые в конце концов слущиваются с поверхности. На тех участках, где эпидермис тонок, он четко подразделяется на пролиферативные единицы, или колонки, по меньшей мере с одной «бессмертной» стволовой клеткой в основании каждой из них. Судьба потомков стволовой клетки отчасти зависит от внешних факторов, еще не вполне выясненных. Скорость пролиферации стволовых клеток регулируется гомеостатически в соответствии с толщиной эпидермиса. В железах, связанных с эпидермисом, например в потовых и молочных железах, тоже имеются стволовые клетки, но обновление клеточной популяции организовано по-иному.