ЗАГАЛЬНА МІКРОБІОЛОГІЯ - Т.П. Пирог - 2004
5. БУДОВА МІКРОБНОЇ КЛІТИНИ
5.1. КЛІТИННІ СТІНКИ МІКРООРГАНІЗМІВ
5.1.1. Поверхневі структури клітинної стінки бактерій
Джгутики і рухливість. За здатністю переміщуватися всі бактерії поділяються на рухливі та нерухливі. У більшості бактерій здатність рухатися зумовлена наявністю джгутиків. Рухатися без джгутиків можуть ковзні бактерії (до них належать міксобактерії, ціанобактерії) та спірохети.
Розміщення джгутиків у рухливих еубактерій є ознакою, характерною для певних груп, тому вона має таксономічне значення. У паличкоподібних бактерій джгутики можуть бути розміщені полярно або латерально (моно- і біполярне розміщення) (рис. 5.1). Серед бактерій з монополярним джгутикуваниям лише деякі мають один, але товстий джгутик (монотрихи) (Vibrio). БІЛЬШІСТЬ бактерій є політрихами. Монополярне-політрихальне розміщення джгутиків називається також лофотрихальним (Pseudomonas. Chromatlum), а біполярне-політрихальне — амфітрихальним (Spirillum). При перитрихальному розміщенні (ентеробактерії, бацили) джгутики розміщуються по боках клітини або по всій поверхні.
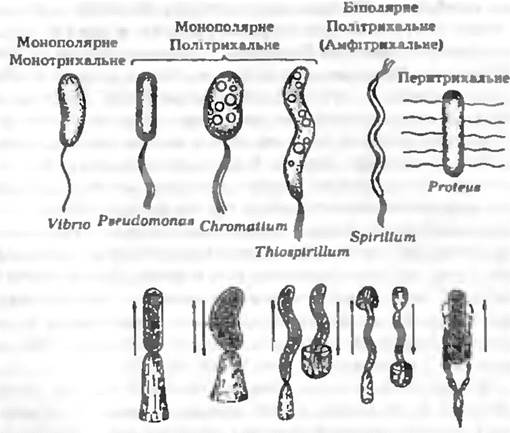
Рис. 5.1. Основні типи джгутикування те тяги руху бактерія
Джгутики являють собою спірально закручені нитки. У різних бактерій вони розрізняються за товщиною (12-18 ям), довжиною (до 20 мкм), а також за довжиною та амплітудою аитка. Нитки джгутиків складаються із специфічного білка флагеліну. Флагелін має молекулярну масу до 40 000.
За функціями флагелін часто порівнюють з міозином м'язових тканин (скорочувальним білком), який забезпечує рух. Але на відміну від більшості скорочувальних білків, функція яких пов’язана з гідролізом АТФ (тобто для забезпечення руху використовується енергія гідролізу АТФ). базальне тіло повертається навколо своєї осі за рахунок енергії, що генерується протон- рушійною силою. Джгутик складається з трьох частин: спіральної нитки, «крюка» поблизу поверхні клітини і базального тільця. За допомогою базального тільця джгутик закріплюється в плазматичній мембрані та клітинній стінці.
Фімбрії та тлі. Поверхня деяких бактерій покрита великою кількістю (від 10 до кількох тисяч) довгих тонких прямих ниток завтовшки 3-25 нм та завдовжки до 12 мкм. які нязиваються фімбріями. Фімбрії зустрічаються у бактерій, які мають і не мають джгутиків. Одна з перших робіт із вивчення хімічного складу фімбрій (у бактерій Escherichia coll) була здійснена у 1960 р.
А. Брінтоном. Експерименти показали, що до складу фімбрій входять пектини — вуглеводзв’язу вальні білки. Залежно від вуглеводної специфічності дентинів існує кілька типів фімбрій. Тяк, лектини фімбрій першого типу є манозоспецифічними. G-фімбрій — специфічними до N-ацетилглюкозаміну. В амінокислотному складі фімбрій першого типу переважають дикарбо- нові та аліфатичні амінокислоти, у той час як сірковмісні амінокислоти присутні у слідових кількостях. У складі фімбріальних лектинів виявлено до 40-50 % гідрофобних амінокислот (пролін, аланін, валін, лейцин, ізолейцин, фенілаланін). Незважаючи на те що фімбрії першого типу утворюються великою кількістю ентеробактерій, амінокислотний склад цих білків встановлений тільки для деяких штамів. Молекулярна маса лектинів, з яких складаються фімбрії, становить близько 16 000-25 000.
Крім фімбрій, клітини багатьох бактерій містять статеві пілі (F-пілі). їх не більше однієї-двох на клітину. Пілі мають вигляд порожніх всередині білкових трубочок завдовжки від 0,5 до 10 мкм. За допомогою статевих пілей чоловіча клітина прикріплюється до жіночої, утворюючи кон’югаційний тунель, по якому відбувається передача ДНК від донора до реципієнта.
Таксиси. Таксиси (від грец. taxis — розміщення) бувають позитивними або негативними, залежно від руху бактерії до фактора чи від нього. Є кілька типів таксисів.
Хемотаксис — рух, який зумовлюється хімічними речовинами. За здатністю індукувати (тобто викликати, зумовлювати) позитивний чи негативний хемотаксис розрізняють дві групи речовин: атрактанти — речовини, що зумовлюють скупчення клітин у ділянці вищої концентрації сполуки; репеленти — речовини, що зумовлюють скупчення клітин у ділянці вищої концентратів. Слід зазначити, шо не всі сполуки, які використовуються мікроорганізмами як поживні речовини, є атрактантами.
Аеротаксис — рух бактерій до (від) молекулярного кисню. У рухливих бактерій тип метаболізму (аеробний чи анаеробний) можна визначити за аеротаксичним рухом і скупченням клітин на певних відстанях від накривного скла. При цьому строго анаеробні бактерії будуть розміщуватися в центрі скла, аероби — біля його країв чи біля бульбашок повітря, факультативні анаероби — між аеробами та анаеробами.
Фототаксис — рух бактерій, зумовлений світловою енергією Так, фототрофні бактерії, яким для одержання енергії необхідне світло, в результаті фототаксису скупчуються в освітленому місці. Якщо витримати в темноті препарат, у якому суспензія клітин Chromatium рівномірно розподілена під накривним склом, а потім спрямувати на нього промінь світла, то бактерії накопичаться в ділянці світлової плями.
Магнітотаксис — рух бактерій за силовими лініями магнітного поля Землі або магніту, зумовлений наявністю в спеціальних гранулах магнітосомах великої кількості заліза (до 0,4 % сухої речовини) у вигляді феромагнітного окису. Магнітосоми розміщені біля місць прикріплення джгутиків.
Термотаксис — рух бактерій, зумовлений джерелом тепла.
Віскозитаксис — рух бактерій у напрямку збільшення чи зниження в’язкості розчину. Наприклад, для спірохет-паразитів людини й тварин, які переміщуються до поверхні слизових оболонок, ця властивість має пристосувальний характер. Механізм цього процесу поки що не встановлений.
Капсула та слизовий шар. Капсула розміщена поверх клітинної стінки. Її можна побачити під світловим мікроскопом, якщо обробити препарат такими барвниками, як нігрозин, конто червоний і китайська туш. які в капсулу не проникають. При цьому маємо як би негативне контрастування: світла капсула виділяється на темному фоні.
Розрізняють мікрокапсули завтовшки 0,2 мкм. Мікрокапсули невидимі у світловому мікроскопі, їх можна виявити тільки імунологічно (за набуханням при змішуванні із специфічними антитілами). Макрокапсула завтовшки більш як 0,2 мкм добре видима у світловому мікроскопі. Слизовий шар за товщиною в багато разів перевищує розміри клітини. Являє собою гідратовину в'язку мису, шо накопичується на поверхні клітини.
Капсулу легко відокремити від клітини механічно, наприклад. центрифугуванням або вилученням у вигляді водних, буферних чи слабколужних розчинів.
За хімічним складом капсули поділяються на:
капсули полісахаридної природи, що складаються з гомополісахаридів (побудовані з одного й того самого моносахариду, наприклад, у Leuconostoc mesenteroides — з глюкози, у бактерій роду Klebsiella — а галактози); гетерополісахаридів (побудовані з різних моносахаридних залишків, наприклад, у Pseudo monas aeruginosa — із залишків глюкози, галактози, манози, рамнози, глюкуронової кислоти);
капсули, що складаються з поліпептидів і полісахаридів, наприклад, у Bacillus megaterium.
Капсулу можна розглядати як пристосувальне утворення у сапрофітних і патогенних бактерій. Утворення капсули стимулюється присутністю живої тканини для патогеиних мікробів (паличка сибірки), наявністю вуглеводів і низькою температурою (тифозна паличка), наявністю сахарози (азотобактер). Полісахариди капсули деяких бактерій є антигенами, вони здатні також сприяти вірулентності бактерій (так, капсульні штами пневмококів спричиняють пневмонію у білих мишей, а декапсульовані втрачають цю здатність).
Капсула утримується на поверхні клітинної стінки за рахунок як іонних, так і ковалентних зв'язків.
Полісахариди, що утворюють капсулу, належать до екзо- полісахаридів (ЕПС). У біотехнології мікробних полісахаридів їх називають капсульними, а полісахариди, що виділяються в культуральиу рідину, — екзополісахаридами.
Здатність до синтезу екзополісахаридів притаманна багатьом мікроорганізмам — представникам різних фізіологічних і таксономічних груп. Синтезуються ЕПС грибами (Aureobasldium pullulans — полісахарид пулулан, Sclerotium rolfsil — склероглюкан), дріжджами (Cryptococcus laurentii, Hansenuiа), бактеріями. Серед бактеріальних продуцентів ЕПС є фітопатогенні бактерії (Xanthomonae campestris, Pseudomonas, Erwinia), азотфіксувальні (Azotobacter beijerinckia), метилотрофні (Mcthylocystis parous, Methylomonasmucosa). Серед ЕПС, як і серед капсульних полісахаридів, є гомо- та гетерополісахариди. Розрізняють також нейтральні ЕПС (складаються тільки з залишків моносахаридів), кислі ЕПС (містять залишки уронових, піровиноградної та інших кислот), лужні ЕПС (містять залишки аміноцукрів).