ЗАГАЛЬНА МІКРОБІОЛОГІЯ - Т.П. Пирог - 2004
5. БУДОВА МІКРОБНОЇ КЛІТИНИ
5.1. КЛІТИННІ СТІНКИ МІКРООРГАНІЗМІВ
5.1.2. Будова і хімічний склад клітинних стінок прокаріот
Хімічний склад клітинних стінок мікроорганізмів спочатку привертав увагу дослідників у галузі систематики. Основою
тому були дані про якісні відмінності у складі клітинних стінок між еукаріотними та прокаріотними мікроорганізмами, а також серед прокаріотів — між грам позитивними та грамвегативними бактеріями (табл. 5.1).
Таблиця 6.1
Хімічний склад клітинних стінок мікроорганізмів
Полімери |
Еукаріоти |
Прокаріоти (бактерії) |
|
Грампозитивні |
Грамнегативні |
||
Пептидоглікан Тейхоєва кислота Ліпополісахарид Ліпопротеїд Білок Полісахарид |
- - - + + + |
+ + - - + + |
+ - + - + + |
Фарбування за Грамом. Диференційоване фарбування бактерій генціанвіолетом було запропоновано у 1884 р. датським фармакологом Г.Х. Грамом. У мікробіологи забарвлення за Грамом є важливою таксономічною ознакою, з якою корелюють інші властивості бактерій. Суть методу полягає в тому, що при фарбуванні бактерій генціанвіолетом (кристалвіолетом, метилвіолетом) фарба з йодом утворюють сполуку, що утримується клітинами при обробці їх спиртом. Такі бактерії забарвлені в синьо-фіолетовий колір і їх називають грампозитивними.
Бактерії, які знебарвлюються при обробці спиртом, називаються грамнегативними. Їх потім дофарбовують контрастною фарбою (фуксином).
У 1978 р. Н.Е. Гіббонс та Р.Г.Е. Муррей запропонували грамнегативні істинні бактерії (еубактерії) виділити у відділ Грацилікутних (Gracilicutes), а грампозитивні — у відділ Фірмікутних (FirmicutPK). Але терміни «фірмікутні", «грацилікуті" бактерії не мають широкого використання в мікробіології,
Пептидоглікан (глікопептид, мукопептид, муреїн). Основним компонентом клітинної стінки бактерій є пептидоглікан (глікопептид, мукопептид, муреїн). Пептидоглікан виявлено тільки у прокаріот. Винятком є еубактерії, що не мають клітинної стінки (мікоплазми. L-форми) та ерхебактерії — деякі метаноутворювальні та галофіли (Halobactehum, Hatococcuа). Для галофільних бактерій наявність міцної клітинної стінки не обов’язкова, оскільки вміст їх клітин є ізоосмотичним з навколишнім середовищем.
Специфічний гетереполімер пептидоглікан складається: із залишків N-ацетилглюкозаміну та N-ацетилмурамової кислоти, з’єднаних між собою β-1.4-глікозидними зв’язками. N-ацетилглюкозаміну є похідною сполукою глюкози, в якій гідроксильна група при другому атомі вуглецю зміщена на аміногрупу. N-ацетилмурамова кислота — це ефір N-ацетилглюкозаміну та D-молочної кислоти (рис. 5.2); із діамінокислот, з яких найчастіше зустрічаються мезодіамінопімелінова кислота. LL-діамінопімелінова кислота, лізин, ориітин. Наявність таких амінокислот з двома аміногрупами має принципове значення для просторової організації пептидоглікану. Вони забезпечують утворення двох пептидних зв’язків між пептидними угрупованнями в молекулі; з інших амінокислот (D- та L аланін, D-глутамінова кислота, Ζ-серин, гліцин).
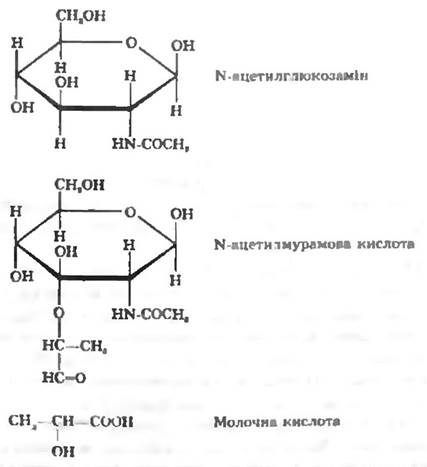
Рис. 5.2. Основні складові гетерополімеру пептидоглікану
Фрагмент пептидоглікану показано на рис. 5.3. За допомогою пептидних місточків гетерополімерні ланцюги зв’язані між собою в мішкоподібну гігантську молекулу — муреїновий мішок (муреїнова сітка).
Муреїновий мішок виконує функцію опорного каркаса клітинної стінки. За будовою цього каркаса, а також за вмістом інших речовин у клітинній стінці грампозитивні бактерії відрізняються від грамнегативних.
Рис. 3.3. Структура пептилоглікану
Клітинна стінка грам позитивних бактерій. У грам позитивних бактерій частка муреїнової сітки становить 30-70 % сухої маси клітинної стінки (завтовшки 40 шарів). Замість мезодіамінопімелінової кислоти часто міститься LL-діамінопімелінова кислота або лізин. У клітинній стійці грампозитивних бактерій полісахариди, якщо вони є, зв’язані між собою ковалентно. Вміст ліпідів і білків невисокий. У білках клітинних стінок грампозитивних бактерій набір амінокислот менший (4-12), ніж у грамнегативних (містяться практично всі амінокислоти, з яких складаються білки).
Харектерною особливістю грампозитивних бактерій є наявність у клітинній стінці тейхоєвих кислот. Тейхоєві кислоти — це ланцюги, які складаються з 8-50 залишків гліцерину чи рибітолу, зв'язаних між собою фосфатними місточками. У молекулі тейхоєвої кислоти поліол може містити моносахариди як замісники. Деякі з тейхоєвих кислот містять еритритол чи меніт. Припускається, що тейхоєві кислоти зв’язані з муреїном через фосфат за типом аміду. У складі тейхоєвих кислот деяких грампозитивних бактерій містяться жирні кислоти, які утворюють ефірні зв’язки з гліцериновими залишками. їх називають ліпотейхоєвиєми кислотами. Тейхоєві кислоти містяться в клітинах у значних кількостях. У деяких бактерій вони становлять більше половини маси клітинної стінки.
Функції тейхоєвих кислот:
фосфатні групи тейхоєвих кислот с місцем зв'язування катіонів магнію, необхідного для багатьох ензиматичних і фізико- хімічних процесів, що проходять на цитоплазматичній мембрані;
тейхоєві кислоти беруть участь у регуляції активності автолітичиих ферментів;
було показано, що цукрові компоненти тейхоєвих кислот с відповідальними за зв’язування фатів з клітинною стінкою. Якщо тейхоєва кислота а будь-яких причин втрачає глікозильні замісники, то бактеріальна клітина стає фато резистентною (фагостійкою);
ліпотейхоєві кислоти беруть участь в імунологічних реакціях.
Клітинна стінка грамнегативних бактерій. У грам негативних бактерій муреїнова сітка с одношаровою і становить менше 10 % сухої маси клітинної стінки. Муреїн містить тільки мезодіамінопімелінову кислоту і не містить лізину. У складі клітинних стінок грамнегативних бактерій тейхоєві кислоти не виявлено.
У всіх грамнегативних бактерій зверху одношарового чи найбільше двошарового муреїнового мішка розміщується зовні штаб шар клітинної стінки. Це так звана зовнішня мембрана, що складається з білків, фосфоліпідів і ліпополісахаридів (ЛПС) (рис. 5.4).
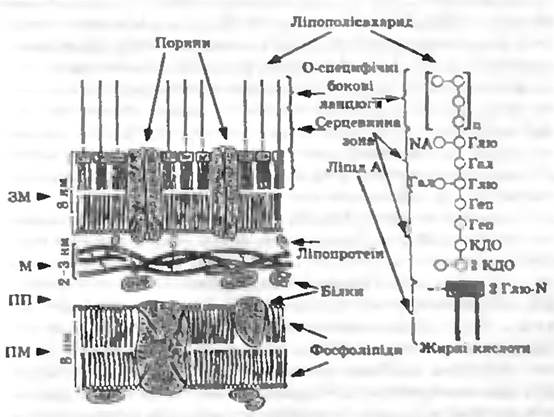
Рис. 5.4. Модель будови клітинної стінки грамнегативних бактерій
З муреїном, очевидно, ковалентно через діамінопімелінову кислоту зв'язані ліпопротеїни. Вони орієнтовані своїми ліпофільними кінцями назовні і таким чином закріплені в ліпофільному подвійному шарі (завдяки гідрофобній взаємодії). У цьому ж шарі містяться фосфоліпіди та гідрофобні кінці ліпополісахаридів. Гідрофільні кінці ЛПС орієнтовані назовні.
Ліпополісахариди — складні молекули a молекулярною масою більш як 10 000. Вони складаються а трьох частин — ліпіду А. ядра (кор, серцевинна зона) та О специфічного бокового ланцюга. ЛПС Salmonella typhimurium та інших ентеробактерій досліджені досить повно.
Ліпід А складається з глюкозаміндисахариду, до гідроксильних груп якого ефірними зв’язками приєднані жирні кислоти (С12. C14. С16). Ця частина молекули має гідрофобні властивості. Далі міститься R-серцевинна зона — трисахарид, що складається з трьох залишків 2-кето-3-дезоксноктонової кислоти (КДО) і який зв’язаний також з фосфоетаноламіном. Далі йдуть дві молекули гептози і зовнішня серцевинна зона. Остання складається з розгалуженого ланцюга, що містить глюкозу, галактозу і N-ацетилглюкозамін. Ця базова структура є однаковою у всіх сальмонел. До серцевинної зони прилягає О-специфічний боковий ланцюг. Це довгі ланцюги, що складаються з повторюваних олігосахаридів, які можуть містити галактозу, манозу, рамнозу, абеквозу, фукозу та інші моносахариди у послідовності, що варіює від штаму до штаму.
ЛПС набули великого значення в бактеріологічній діагностиці і розпізнаванні епідемій. Виявилось, що збудники різних захворювань відрізняються один від одного О-специфічними боковими ланцюгами. Незначні відмінності в їх складі можуть бути виявлені за допомогою імунологічних методів. За серологічними реакціями у роді Salmonellaвдалося виділити понад тисячу видів і штамів. Є так звані місцеві раси сальмонел, які можна ідентифікувати за імунохімічними особливостями. Це часто дає можливість встановити, де відбулося зараження хворого чи звідки почала поширюватися епідемія. Наприклад, можна встановити, де хворий отримав інфекцію — у південно-американському чи східно-азіатському регіоні.
Функції зовнішньої мембрани. Зовнішня мембрана грамнегативних бактерій виконує не тільки механічні, а й важливі фізіологічні функції. В її подвійний ліпідний шар, що складається з ліпіду А, полісахаридів і фосфоліпідів вбудовані білки, які пронизують цей шар наскрізь. Ці трансмембранні білки називаються поринами. Порини пропускають через мембрану гідрофільні низькомолекулярні речовини (до молекулярної маси близько 6 000).
Зовнішня мембрана прилягає до муреїнового шару і зв’язана з ним ліпопротеїнами. Очевидно, муреїновий шар є проникним для різних сполук. Проміжок між муреїном і плазматичною мембраною називають периплазматичним простором. У ньому містяться ферменти, в тому числі і деполімерази (протеїнази, нуклеази), периферійні білки і так звані зв’язувальні білки. Останні беруть участь у перенесенні деяких субстратів у цитоплазму і є рецепторами хемотаксичних сигналів. Пернплазматичний простір, очевидно, відіграє певну роль в осморегуляції.
Дія лізоциму та пеніциліну. Структура клітинної стінки та муреїну була встановлене у зв’язку з вивченням дії лізоциму та пеніциліну на бактерії. Відкритий англійським мікробіологом А. Флемінгом у 1922 р. лізоцим є бактерицидним ферментом. що міститься в яєчному білку, носовому слизі, в сльозовій рідині. Лізоцим виділили також з бактерій (Е. соli, Strrptomyce») і бактеріофагів. При дії лізоциму на суспензію грампозитивних бактерій спостерігали швидке її просвітлення. Так. Micrococcus luteus лізується (розчиняється) вже у концентрації 1 мкг лізоциму на 1 мл. Для лізису клітин Bacillus megaterium необхідною є концентрація 50 мкг/мл, а грамнегативні бактерії розчиняються тільки за наявності в суспензії ЕДТА.
Лізоцим розриває в муреїні глікозидний зв'язок між першим вуглецевим атомом N-ацетилмурамової кислоти і четвертим вуглецевим атомом N-ацетилглюкозаміну (див. рис. 5.3). При цьому полісахаридні ланцюги розщеплюються до дисахаридних фрагментів. Отже, лізоцим є N-ацетилмурамідазою.
Слід зазначити, що повному руйнуванню бактеріальних клітин можна запобігти, здійснюючи лізне в ізотонічному чи слабкогіпертонічному розчині (0,1 -0,2 М сахарози). У цих умовах під дією лізоциму з клітин утворюються надзвичайно чутливі до осмотичних умов округлі протопласти. У гіпертонічних та ізотонічних розчинах протопласти стабільні, у гіпотонічних — лопаються. Протопластами слід називати тільки такі округлі клітини, в яких немає ніякнх залишків клітинної стінки, тобто не можна виявити ні мурамової кислоти, ні специфічної для клітинної стінки діамінопімеліновоі кислоти. Лізис клітинної стінки не супроводжується порушеннями метаболізму.
Крім лізоциму, є ряд інших ферментів, що руйнують муреїновий каркас, наприклад, муроендопептидази.
Антибіотик пеніцилін діє переважно на грампозитивні бактерії (пневмококи та стафілококи), а також на деякі грамнегативні (гонококи, менінгококи, ентеробактерії), вбиваючи їх. Але бактерицидній дії піддаються тільки клітини, які ростуть. Клітини. що перебувають у стані спокою, залишаються живими. Найцікавіший феномен, який спостерігається під дією пеніциліну. — це поява так званих форм, які утворюються а нормальних бактеріальних клітин у результаті незбалансованого росту в довжину та ширину. При цьому вихідні палички збільшуються в об’ємі в багато разів. Якщо діяти пеніциліном на клітини, що ростуть, у гіпотонічному розчині вони лопаються. В ізо- та гіпертонічних розчинах палички перетворюються на шароподібні утворення, які називаються L-формами або сферо пластами. Від протопластів вони відрізняються тим, що зберігають залишки клітинної стінки. Пеніцилін порушує процес утворення клітинної стінки.