Биологические мембраны - А. Н. Огурцов 2012
Электрогенез биомембран
Механизмы межклеточной сигнализации
Синапсы между нейронами
Синапсы между мотонейронами и клетками скелетных мышц (нейромышечное соединение) (рисунок 154) работают как триггеры (каждый потенциал действия в пресинаптическом нейроне вызывает сокращение клетки мускулатуры), поскольку потенциал действия в пресинаптической клетке стимулирует секрецию такого количества ацетилхолина, которого достаточно для деполяризации мембраны постсинаптической клетки.
Синапсы между нейронами работают иначе. Рассмотрим синапс между сенсорным болевым нейроном и ассоциативным (связывающим, relay) нейроном (рисунок 159).
В этом синапсе один импульс потенциала действия в пресинаптической части аксона сенсорного нейрона стимулирует нейросекрецию такого (небольшого) количества нейротрансмиттерных молекул глутаминовой кислоты в синаптическую щель, что связывание их с глутаминовыми рецепторами на постсинаптической мембране ассоциативного нейрона хоть и приводит к деполяризации этой мембраны, но амплитуда её оказывается ниже порога генерации потенциала действия.
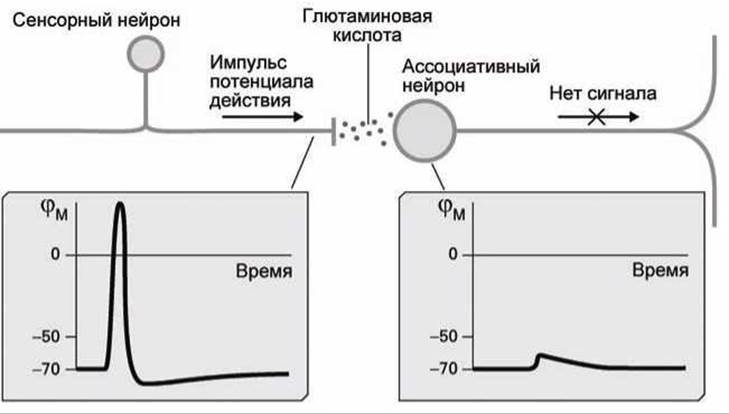
Рисунок 159 - Единичный импульс в сенсорном нейроне не способен стимулировать потенциал действия в ассоциативном нейроне
Поэтому после удаления глутаминовой кислоты из синаптической щели мембранный потенциал постсинаптической мембраны возвращается к значению потенциала покоя.
Если же на дендриты ассоциативного нейрона приходят импульсы с нескольких сенсорных нейронов (рисунок 160), возбуждаемых одновременно, то в результате такого пространственного суммирования "дозы" глутаминовой кислоты суммируются, деполяризация мембраны превышает порог, в постсинаптическом ассоциативном нейроне развивается импульс потенциала действия, и организм чувствует боль.
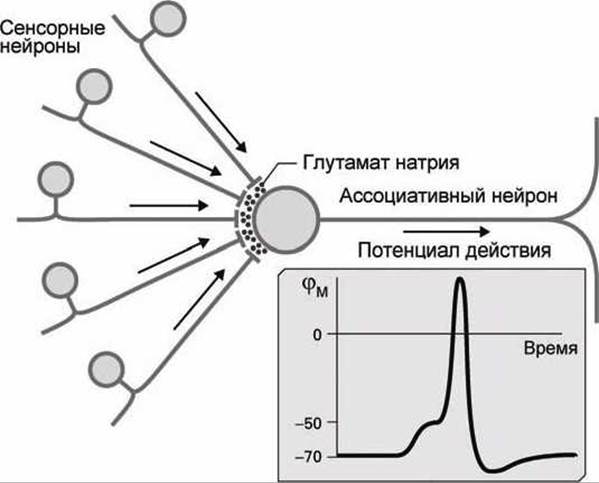
Рисунок 160 - Пространственное суммирование на синапсе
Потенциал действия может быть сгенерирован в постсинаптическом ассоциативном нейроне и в результате инициации одним-единственным пресинаптическим нейроном. Это возможно в случае интенсивного возбуждения рецептора сенсорного нейрона, который выдаёт непрерывную последовательность импульсов с малой скважностью (рисунок 161).
Например, в случае с болевым рецептором (рисунок 137) если уколоть палец иголкой, то этот укол стимулирует только один рецептор в пальце, но интенсивность возбуждения рецептора будет настолько велика, что в рецепторе возбуждается непрерывная последовательность импульсов потенциала действия.
Каждый из этих импульсов стимулирует высвобождение глутамата натрия в синапс, и, если расстояние между отдельными импульсами этой последовательности достаточно мало, происходит процесс временного суммирования деполяризаций, мембранный потенциал достигает порогового значения, и возбуждается потенциал действия, который, распространяясь по аксону ассоциативного нейрона, передаёт информацию о событии в мозг (рисунок 161).
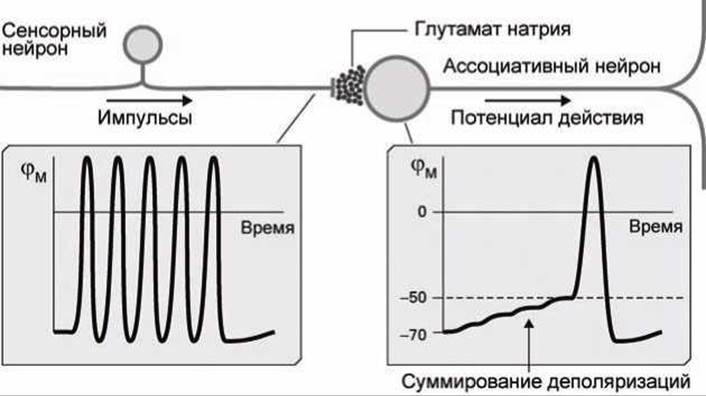
Рисунок 161 - Временное суммирование на синапсе
При суммировании импульсов возбуждения от нескольких пресинаптических нейронов на постсинаптическом нейроне, а в особенности на нейронах головного мозга, возможны и более сложные ситуации, поскольку нейрон различным образом реагирует на нейротрансмиттеры различного типа от различных нейронов.
Рассмотрим, например, ситуацию, при которой ассоциативный нейрон образует шесть синапсов с пресинаптическими нейронами, секретирующими глутамат натрия в синаптические щели, и один синапс с аксоном нейрона, использующим у-аминомасляную кислоту (GABA, рисунок 152) в качестве нейротрансмиттера (рисунок 162).
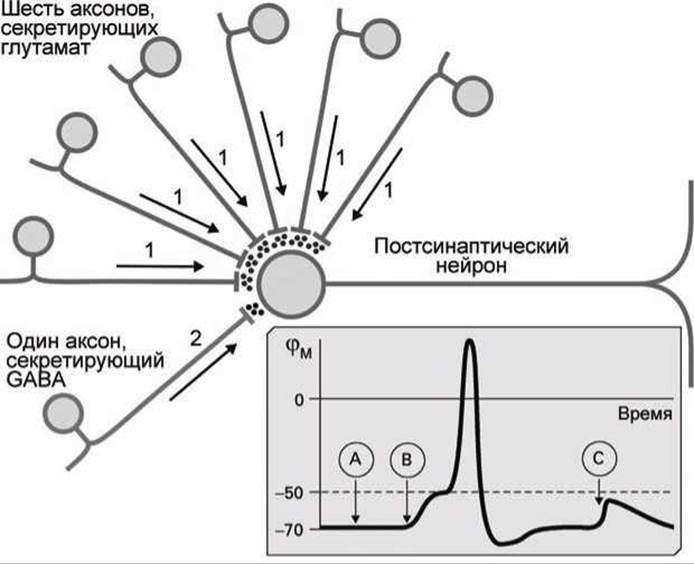
Рисунок 162 - Ингибирование GABA-синапсом
Рецептор у-аминомасляной кислоты является управляемым белковым каналом для ионов Сl-. В нашем примере постсинаптический нейрон имеет рецепторы как для глутамата, так и для GABA.
Рассмотрим, как будет реагировать постсинаптический нейрон при различных комбинациях сигналов, поступающих от пресинаптических нейронов.
В ситуации, обозначенной (А) на рисунке 162, потенциал действия (2) в GABA-секретирующем аксоне высвобождает GABA, и GABA-рецепторы постсинаптического рецептора открываются. Хотя теперь анионы хлора и могут проходить через мембрану, но электрохимические потенциалы ионов СГ равны по обе стороны мембраны и градиент концентрации уравновешен электрическим градиентом. Поэтому открытие GABA-каналов не меняет мембранный потенциал.
В ситуации (В) в синапсы шести глутамат-секретирующих аксонов одновременно приходят импульсы потенциала действия (1). В этом случае в синаптических щелях оказывается достаточно молекул нейротрансмиттеров, чтобы произошла деполяризация мембраны постсинаптического нейрона и генерации в нём потенциала действия.
Рецепторы, связывание нейротрансмиттеров с которыми приводит к генерации потенциала действия, называются возбуждающими рецепторами (excitatory receptors). Возбуждающие рецепторы нейронов являются каналами для ионов Na+. В нашем примере возбуждающими являются глутамат-управляемые рецепторы.
В ситуации (С) возбуждающие импульсы приходят одновременно и от шести глутамат-секретирующих аксонов (1), и от GABA-секретирующего аксона (2). Как и в ситуации (В) такое же количество глутамата натрия секретируется в синаптические щели, и такое же количество ионов натрия проходит через глутамат-управляемые рецепторы, стремясь деполяризовать постсинаптическую мембрану.
Однако, поскольку мембранный потенциал постсинаптической мембраны уже не равен потенциалу покоя, то условие равновесия для ионов хлора нарушено, и ионы С1 проходят через мембрану внутрь постсинаптического нейрона через каналы GABA-рецепторов.
Поток анионов хлора частично нейтрализует избыточный положительный потенциал, который образуется за счёт потока катионов натрия внутрь постсинаптического нейрона через каналы глутаминовых рецепторов. Поэтому деполяризация постсинаптического нерва не достигает критического значения, и потенциал действия не возбуждается.
В нашем примере срабатывание GABA-рецепторов приводит к ингибированию генерации нервного импульса.
Рецепторы, связывание нейротрансмиттеров с которыми приводит к ингибированию потенциала действия, называются ингибирующими рецепторами (inhibitory receptors). Ингибирующие рецепторы нейронов являются каналами для ионов Сl-, входящих в клетку, или для ионов К+, выходящих из клетки. В нашем примере ингибирующими являются GABA-рецепторы.
Успокоительные лекарственные препараты, такие, как валиум, действуют на GABA-рецепторы, облегчая открытие каналов. Те нейроны, на которые подействовал валиум, реже деполяризуются, следовательно, валиум снижает активность нейронов мозга, успокаивая пациента.
Контрольные вопросы и задания
1. Какие межклеточные соединения называются щелевыми?
2. Как устроены щелевые межклеточные соединения?
3. В каких случаях межклеточные соединения должны быть щелевыми?
4. Что такое трансмиттеры?
5. Как различаются трансмиттеры по времени жизни? Приведите примеры.
6. Что такое паракринные трансмиттеры?
7. Какие различают три типа рецепторов для трансмиттеров?
8. С каким трансмиттером связывается мембранный а-адренергический рецептор?
9. С каким трансмиттером связывается мембранный ß-адренергический рецептор?
10. Какие внутриклеточные рецепторы трансмиттеров вы знаете?
11. Какие нейротрансмиттеры вы знаете?
12. Какие существуют два отличия нейросекреции от других секреторных механизмов?
13. В чём состоит отличие рециклирования ацетилхолина из синаптической щели в пресинаптический аксон от рециклирования остальных нейротрансмиттеров?
14. Какие два типа синаптических везикул разделяют в терминалях аксонов?
15. Какое межклеточное соединение называют нейромышечным (или нейромускулярным)?
16. Какие четыре типа ионных каналов и в какой последовательности обеспечивают мышечное сокращение?
17. Из каких субъединиц состоит никотиновый ацетилхолиновый рецептор?
18. Какие конформации субъединиц обеспечивают работу никотинового ацетилхолинового рецептора?
19. Какой нейротрансмиттер секретируется сосудосуживающими нервами?
20. Какой нейротрансмиттер секретируется сосудорасширяющими нервами?
21. Какой фермент чаще всего использует цАМФ для метаболического действия в сигнальных цепях?
22. Какой фермент чаще всего использует цГМФ для метаболического действия в сигнальных цепях?
23. Почему оксид азота является паракринным трансмиттером?
24. Какие выделяют три основные изоформы NO-синтаз?
25. В чём состоит функциональное отличие синапсов между мотонейронами и клетками скелетных мышц от синапсов между двумя нейронами?
26. Каким образом происходит пространственное суммирование нервных импульсов на синапсе между нейронами?
27. Каким образом происходит временное суммирование нервных импульсов на синапсе между нейронами?
28. Какие рецепторы называются возбуждающими?
29. Какие рецепторы называются ингибирующими?
30. В чём сходство и отличие процессов в синапсах возбуждающих и ингибирующих рецепторов с нейроном?