ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 1 - Г.Й. Щербак - 1995
ПІДЦАРСТВО БАГАТОКЛІТИННІ (METAZOA)
РОЗДІЛ СПРАВЖНІ БАГАТОКЛІТИННІ (EUMETAZOA)
ТИП КИШКОВОПОРОЖНИННІ (CNIDARIA, АБО COELENTERATA)
КЛАС КОРАЛОВІ ПОЛІПИ (ANTHOZOA)
Підклас Зоантарії (Zoantharia)
Це поодинокі або колоніальні корали, більшість яких має зовнішній вапняковий (карбонатний) скелет. Відомі також безскелетні форми. Кількість щупалець і септ у га- стральній порожнині, як правило, кратна шести. Щупальця нерозгалужені. Мезоглея тонка.
До цього підкласу належать кілька сучасних рядів, серед яких найбільш відомі: Склерактинії, або Мадрепорові корали (Scleractinia, або Madreporaria), Актинії (Actiniaгіа), а також вимерлі форми; найбільш численні у викопних рештках представники рядів Табуляти (Tabulate) та Ругози, або Чотирипроменеві корали (Rugosa).
Серед усіх рядів зоантарій особливе значення мають склерактинії, або мадрепорові корали (Scleractinia), які є основними будівниками коралових рифів завдяки своєму масивному вапняковому скелету.
Ряд Актинії (Actiniaria)
Це переважно поодинокі безскелетні поліпи, які нагадують фантастичні квіти й тому називаються ще морськими анемонами. Вони мають циліндричну форму з добре розвиненим ротовим диском,на одному кінці й розширеною основою (підошвою) на протилежному (рис. 117). Епідерма підошви виділяє слиз, яким актинії тимчасово прикріплюються до субстрату. Навколо ротового диска концентричними колами розташовані 16—1 000 щупалець. На щупальцях містяться батареї жалких клітин. Рот має вигляд щілини, глотка сплющена й містить два сифоног гліфи, биттям їх війок створюються два токи води: один спрямований всередину гастральної порожнини, другий — із гастральної порожнини назовні.
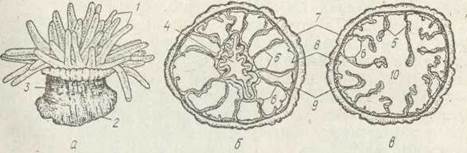
Рис. 117. Ряд Actiniaria — загальний вигляд (а), поперечний розріз на рівні глотки (б) та гастральної порожнини (в):
1 — щупальці; 2 — підошва; 5 —тулуб; 4 — глотка; 5 —септи; 6 — мускульні валки; 7 — епідерма;
8 — гастродерма; 9 — мезоглея; 10 — шлунок
У більшості актиній число септ у гастральній порожнині кратне шести, рідше — восьми або дев'яти.
Більшість актиній — хижаки, живляться ракоподібними, молюсками, навіть дрібними рибами, вбиваючи їх жалкими капсулами щупалець. Але є й види, які живляться найдрібнішими організмами планктону (бактеріями, водоростями, найпростішими), заганяючи їх війками, що містяться на клітинах епідерми щупалець і ротового диска, до травної порожнини.
Розмножуються актинії нестатевим і статевим способами. Нестатеве розмноження рідко відбувається шляхом брунькування. Найчастіше це поздовжній поділ. При цьому спочатку розділяється рот, потім ротовий диск і, нарешті, все тіло актинії. Рідше відбувається поперечний поділ — посередині тіла актинії виростає віночок щупалець, потім верхня, частина відшнуровується й відділяється від нижньої. У верхньої частини відновлюється підошва, а у нижньої — утворюються ротовий диск і глотка.
Крім згаданих способів нестатевого розмноження, є ще один — лацерація, внаслідок якої одразу утворюється кілька дрібних особин. Від підошви дорослої актинії відокремлюється невеличка її ділянка, з якої потім розвиваються нові актинії. Актинії, як і гідри, здатні до регенерації.
Під час статевого розмноження в стінках септ визрівають статеві клітини, що мають ендотермальне походження. Актинії, як правило, роздільностатеві, але трапляються й гермафродити. Запліднення може відбуватися у воді або в гастральній порожнині материнського організму. Плаваючі личинки — планули — розносяться морськими течіями на значні відстані.
Актинії трапляються в усіх морях і океанах — від Арктики до антарктичних вод, але найрізноманітніші вони в тропіках. У Чорному морі мешкає лише два види: Actinothoe clavata та Actinia equina, останній вид знайдено й у Азовському морі.
Більшість видів мешкає в прибережному мілководді, але є й глибоководні форми, деякі з них живуть на максимальних глибинах Світового океану (до 11 км).
Окремі види актиній вступають у мутуалістичні відносини з іншими тваринами. Класичний приклад —це співіснування раків-самітників і актиній Adamsia. Актинія оселяється на черепашці молюска, в якій живе рак-самітник. Така асоціація є корисною для обох видів. Актинія одержує залишки їжі рака-самітника, набуває рухливості, а рак-самітник перебуває під надійним захистом щупалець актинії.
Один із середземноморських крабів постійно тримає в клешнях двох актиній для захисту від ворогів і захоплення їжі, яку актинії ловлять своїми щупальцями, а краб відбирає.
Має місце також симбіоз велетенських актиній Stoichactis, що живуть на західному узбережжі Австралії, з рибками-амфіприонами, які перебувають серед щупалець актинії, ховаючись від ворогів. У щупальцях виробляється особлива речовина, що гальмує вистрілювання жалких капсул. Ця речовина поступово накопичується в слизі, що вкриває тіло рибок, тому вони після перших, дуже обережних контактів із актинією поступово стають несприйнятливими до її отрути. Актинії одержують залишки їжі рибок, крім того, риби постійним биттям плавців сприяють поліпшенню газообміну актинії.
Ряд Склерактинії, або Мадрепорові корали (Scleractinia, або Madreporaria)
Склерактинії — найчисленніша група коралів, яка налічує, за сучасними відомостями, близько 550 видів. Це переважно колоніальні, рідше поодинокі корали, що мають вапняковий скелет ектодермального походження. Скелет у склерактиній суцільний, масивний.
У поодиноких поліпів скелет утворюється спочатку на підошві, епідерма якої складається з особливих клітин — калікобластів (склеробластів). На поверхні цих клітин виділяються наитонші хітинові ниточки, які, переплітаючись між собою, утворюють дуже ніжну щільну сіточку. На ній і відкладаються кристалики вапна (СаСО3), які зливаються потім у суцільний скелет, що постійно наростає. Таким чином утворюється підошовна пластинка, з якої розвиваються скелетні перетинки — склеросепти, що вростають у м'які тканини поліпа. У периферійній області поліпа від підошви відходить невисока чашечка-гесо, яка оточує його нижню частину. Так виникає кораліт — скелет окремого поліпа. М'які тканини опиняються всередині кораліта, вони щільно прилягають до нього та повторюють його рельєф (рис. 118).
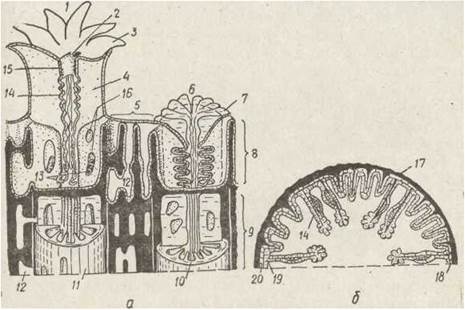
Рис. 118. Схема будови склерактиній — ділянка колонії (а), поперечний зріз окремого поліпа (б): 1 — розправлений поліп; 2 — рот; 3—щупальця; 4 — м'яка септа; 5 — ценосарка; 6 — втягнутий поліп; 7 — склеросепта; 8 — м'яке тіло; 9 — відмерлі поліпи (голий скелет); 10— денце; 11 — кораліт; 12 — порожнина ценосарка; 13 — аконції; 14 — мезентеріальні нитки; 15— глотка; 16 — отвори у склеросептах; 17— склеротика; 18— мезоглея; 19 — гастродерма; 20 — епідерма
У колонії склерактиній розрізняють окремі поліпи (зоріди) та ценосарк. Тіло зооїда складається з нижньої, зануреної в скелетну чашечку кораліта частини, та верхньої, що піднімається над коралітом. Під час подразнення корала верхня частина поліпа вгортається всередину нижньої. Верхня частина тіла поліпа розправляється за рахунок нагнітання води через рот. Сифоногліфів у склерактиній немає. Рот і глотка в них виконують функцію клапана, що запобігає відтоку води з гастральної порожнини, й результаті чого тіло поліпа опирається на замкнену всередині нього воду — своєрідний гідроскелет.
На підошві в зооїдів колонії утворюються скелетні денця, що відокремлюють скелети живих поліпів від скелетів попередніх поколінь, які лежать під ними.
М'яке тіло колонії розташоване на поверхні загального скелета — політика. Набуваючи його форми, воно вкриває скелет у вигляді сплющеного мішка, стінки якого складаються з епідерми, тонкої мезоглеї та гастродерми. Нижній бік мішка щільно обгортає поверхню скелета, заходячи вглиб його порожнин. Підошви окремих поліпів з'єднані між собою в суцільний покрив, епідерма (калікобласти) якого на нижньому боці виділяє вапно, що заповнює простір між поліпами й утворює скелет ценосарка. Порожнини ценосарка сполучаються одна з одною та з гастральними порожнинами отворами або трубочками; всі порожнини вистелені гастродермою (див. рис. 118).
Отже, живі поліпи містяться лише в поверхневому шарі колонії, а вся товща поліпняка утворена скелетами минулих поколінь. Під час росту колонії в процесі поновлення поліпів вони створюють над старими днищами нові, причому м'які тканини залишаються лише в поверхневій частині кораліта, а стовпчик із днищ тягнеться вглиб скелета. За об'ємом і масою скелет багаторазово переважає живі тканини, які ніби розтікаються тоненьким шаром по поверхні масивного поліпняка.
За формою колонії склерактиній надзвичайно різноманітні, причому будова колонії залежить переважно від характеру руху водних мас. Один і той же вид коралів у більш спокійній воді будує кулястий поліпняк, в менш спокійній — гіллястий, в бурхливій — плескатий, що стелеться по дну (рис. 119).
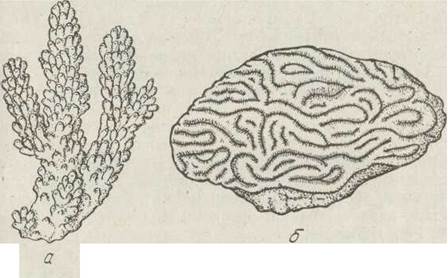
Рис. 119. Ряд Scleractinia: а — Аеrороrе; б — Leptorla
Тривалий час особливості живлення, дихання та процеси, -що зумовлюють утворення могутнього скелета, не вивчалися. За аналогією з іншими кишковопорожнинними склерактиній вважали хижаками. Нині з'ясовано, що живлення склерактиній відбувається трьома способами.
1. Склерактинії використовують ту їжу, яку приносять їм морські течії: дрібні планктонні організми, яйця, цисти, органічній часточки тваринного та рослинного походження (псевдапланктон). Важливу роль в уловлюванні та поглинанні їжі відіграють війки, що вкривають оральний диск, а також слиз. Поліпи випускають з рота слизисті тяжі, які витягуються між щупальцями. Під дією сильних токів води, що виникають внаслідок биття війок, слиз розпушується, утворюючи сітку клейких ниток, до якої прилипають харчові часточки. Періодично слиз разом із осілими на ньому часточками їжі втягується до рота. Цей механізм поліпи використовують не тільки для фільтрації, а й для хижого живлення. Рухлива здобич вбивається жалкими клітинами щупалець, потрапляє в слизові сітки, приклеюється до них і заковтується разом із слизом.
Нещодавно відкрито ще один спосіб живлення склерактиній— перетравлення осілих на поверхні тіла харчових часточок ферментами, що секретуються всією поверхнею епідерми, її клітини здатні також захоплювати бактерій, що осідають на їх поверхні. Проте тропічні води бідні на планктон, тому склерактинії в такий спосіб свої потреби не задовольняють.
2. Результати дослідів із радіоізотопами свідчать, що склерактинії здатні засвоювати з води через покриви органічні речовини (амінокислоти, пептиди тощо), вилучаючи їх із навколишнього середовища. Епідерма поліпів має надзвичайно складну будову. Вона вкрита мікроворсинками, впинаннями мембрани, що збільшує контакт поверхні епідермальних клітин із зовнішнім середовищем, полегшуючи засвоєння з води молекул розчинених у ній органічних речовин. Великі колоїдні частки їжі засвоюються шляхом піноцитозу, а дрібні молекули — шляхом активного ферментного транспорту. Це було доведено в дослідах з коралами роду.
3. Склерактинії одержують поживні речовини також за допомогою симбіонтів — зооксантел. Це вид динофлагелят Symbiodinium microadriaticum.
Симбіодініум не трапляється у вільному стані, проте він здатний вступати в симбіотичні відносини з багатьма безхребетними — кораловими поліпами, губками, медузами, війчастими червами, молюсками і навіть найпростішими (інфузоріями, радіоляріями, форамініферами). Сим- біодініуми містяться в клітинах гастродерми або в міжклітинниках. Вирости клітин гастродерми хазяїна утворюють своєрідну капсулу навколо зооксантели. Від клітини хазяїна її відокремлює складної будови тришарова оболонка, що містить клітковину. Ця оболонка надійно ізолює симбіонта від атаки лізосом хазяїна, запобігаючи його перетравленню. Разом із тим вона проникна для метаболітів у процесі обміну симбіонта з його клітинами.
За допомогою мічених за С14 молекул СO2 виявлено, що зооксантели синтезують органічні речовини, які засвоюють корали. У процесі фотосинтезу в зооксантелах утворюються переважно жири, але вони не накопичуються в зооксантелах, а переходять у клітини поліпа. Жир — це основний енергетичний матеріал поліпів, який є також основною запасною речовиною. Зворотний рух метаболітів із клітин хазяїна в клітини симбіонта полягає перш за все у споживанні зооксантелами біогенних речовин — солей азоту та фосфору. Зооксантели можуть також засвоювати з клітин хазяїна складні органічні молекули типу нуклеотидів і поліпептидів.
Склерактинії дихають киснем, розчиненим у воді, але види поліпів, що мають симбіотичні водорості, одержують його безпосередньо в товщі тканин.
Симбіодініум знаходить у коралах притулок і захист від ворогів. Він використовує для фотосинтезу вуглекислий газ, який виділяють поліпи в результаті дихання, а також утилізує азот і фосфор, що містяться в продуктах метаболізму коралів.
Ті види коралів, які мають симбіотичні водорості, називаються герматипними, а ті, що не мають, — агерматипними. Герматипні корали можуть довго жити в чистій воді без усякої їжі, лише за рахунок симбіонтів. Якщо їх штучно позбавити водоростей (утримуючи деякий час у темряві), вони припиняють ріст і розмноження, а потім, незважаючи на достатню кількість їжі, гинуть.
Крім зазначених переваг у живленні та диханні, герматипні корали значно швидше (майже в десять разів), ніж агерматипні, можуть нарощувати свій скелет. Тому серед рифоутворювачів переважають саме герматипні види. Механізм нарощування вапнякового скелета такий. У воді завжди є невеличка кількість СО2 та іонів
Са24-. Під час розчинення СО2 у воді незначна його частина (близько 1 %) вступає в реакцію з водою, утворюючи вугільну кислоту:
![]()
Вугільна кислота — нестійка сполука, вона може існувати лише у водному розчині, причому розпадається на іони:
![]()
Під час взаємодії іонів Са2+ і НСО3- утворюється вуглекальцієва сіль (гідрокарбонат кальцію):
![]()
Ця речовина розчинна у воді, але нестійка й легко перетворюється на нерозчинну сіль — карбонат кальцію - (вапно):
![]()
У разі надлишку СО2 реакція зсувається ліворуч, тобто в бік утворення розчинного гідрокарбонату, а внаслідок нестачі СО2 — праворуч, що супроводжується випадінням осаду вапна. Протягом світлого часу доби симбіодініуми в процесі фотосинтезу безперервно поглинають із тканин коралів ССb, завдяки цьому вміст його у внутрішньому середовищі організму коралів знижується, що сприяє зсуванню хімічної реакції в бік осадження вапна.
Фотосинтез у симбіодініума відбувається найбільш - ефективно в умовах інтенсивного освітлення. Тому герматипні корали оселяються на незначній глибині, що не перевищує 50 м, утворюючи рясні зарості — рифи. Кораловими рифами називаються масові поселення морських організмів, що мають твердий вапняковий скелет. Основу такого рифу становлять мадрепорові корали (склерактинії), але існує багато організмів, які теж мають вапняковий скелет і беруть участь в утворенні рифу — вапнякові водорості, гідрокорали, сидячі поліхети, що живуть у вапнякових трубках, молюски, вусоногі раки. Проте чільну роль в утворенні рифу відіграють все ж таки склерактинії.
Корали-рифоутворювачі потребують досить високої й постійної температури води (не нижче 20°С), нормальної океанічної солоності (35) і освітлення. Тому коралові" рифи поширені винятково в тропіках.
Розрізняють берегові рифи, що оточують береги островів або материків, бар'єрні рифи, які тягнуться вздовж берега на деякій відстані від нього, атоли — кільцеподібні коралові острови. Є також коралові банки, що виникають на мілинах, і густі коралові поселення в лагунах і бухтах.
Походження коралових споруд здавна цікавило зоологів і геологів. У різні часи було висунуто багато гіпотез, але найбільш переконливою залишається теорія, створена ще Ч. Дарвіном у 1842 р. На його думку, першою стадією утворення коралових островів є облямовуючий береговий риф. Корали використовують у цьому випадку береги островів як субстрат для будівництва рифу. Якщо в результаті тектонічних явищ у земній корі дно моря починає опускатися, й острів поступово занурюється у воду, береговий риф, який облямовує, з тією ж швидкістю росте вгору та вздовж зовнішнього краю, де більш чиста й багата на кисень вода. Корали внутрішнього краю., оберненого в бік острова, поступово відмирають, і береговий риф перетворюється на бар'єрний, який все далі відступає від берега. Коли острів повністю заглиблюється під воду, на його місці утворюється лагуна, а бар'єрний риф перетворюється на атол.
У Індійському та Тихому океанах розкидані сотні таких атолів діаметром від 1 до 160 км, але більшість з них має діаметр 3—30 км. Загальна площа сучасних рифів становить близько 600 тис. км2.
Кораловий риф — це своєрідна екосистема, до складу якої входять не тільки організми-рифоутворювачі, а й величезна кількість інших тварин і рослин. Автотрофні організми (в основному симбіодініум) синтезують органічну речовину, тварини її споживають, поїдаючи м'які тканини поліпів. Серед них переважають молюски, поліхети, морські зірки, краби, риби. Багато тварин, такі, як сидячі в трубках поліхети, вусоногі раки, голотурії, знаходять тут притулок, а живляться завислими у воді органічними рештками; інші, такі як нематоди, морські їжаки, офіури, живляться детритом. Є й хижаки, які вживають в їжу інших мешканців рифів.
Характерна особливість коралових рифів полягає в їх надзвичайно високій біологічній продуктивності. Автотрофна фотосинтетична продукція в екосистемі-рифу становить від 50 до 300 г сирої біомаси на 1 м2 за добу.
Отже, екосистеми коралових рифів одні з найбільш зрілих і ефективно функціонуючих біогеоценозів нашої планети. Завдяки надзвичайно високій продуктивності й здатності до біофільтрації сучасні рифи відіграють важливу роль у глобальних біогеохімічних процесах, а також у динаміці Світового океану.
Протягом тисячоліть коралові рифи становили основу життя цілих народів, мешканців островів Океанії. Ці острови складаються з коралового вапняка. Завдяки багатовіковому досвіду полінезійці, меланезійці та мікронезійці користувалися всіма багатствами рифу, не завдаючи шкоди його існуванню.
Для коралових рифів згубними є опріснення (наприклад, тропічні зливи під час відливу), осушення, урагани — після таких катастроф риф відновлюється протягом десятків років. Коралами живляться деякі риби, краби, морські зірки та інші організми, але їх кількість регулюється іншими хижими тваринами. У разі порушення біологічної рівноваги внаслідок забруднення води або посиленого вилову окремих видів тварин можуть надмірно розмножитися певні види хижаків, які живляться коралами, що призводить до загибелі цілих масивів коралів. Така катастрофа загрожувала в останні роки Великому Бар'єрному Рифу, що розташований біля берегів Австралії, через надмірне розмноження морської зірки «терновий вінець» (Acanthaster planci), якби не своєчасні заходи уряду. Отже, екосистема коралового рифу дуже своєрідна й уразлива, вона здатна існувати лише в дуже обмеженому діапазоні зовнішніх умов, тому потребує захисту.